| |
|
本文对应知识领域
|
第二章 植物的水分生理(五)
作者:未知
申领版权
2010年10月08日
共有 4078 次访问
【添加到收藏夹】
【我要附加题目】 |
|
受欢迎度:
第五节 植物体内水分向地上部分运输
一、水分运输的途径和速度
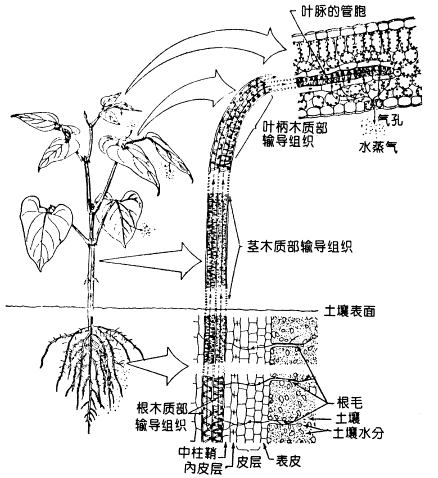
在土壤—植物—大气连续体中,水分运输的具体途径是:土壤→根毛→皮层→内皮层→中柱鞘→根的导管或管胞→茎的导管→叶柄导管→叶脉导管→叶肉细胞→叶细胞间隙→气孔下腔→气孔→大气(图2-15)。水分在植物根、茎、叶内的运输既有质外体运输,又有共质体运输。其中一部分是在活细胞中进行的短距离径向运输(如从皮层→根中柱,叶脉→叶肉细胞)。共质体运输虽只有几毫米,但水分通过原生质体部分的阻力大,运输速度很慢,一般只有10-3cm·h-1。另一部分是通过导管或管胞的长距离质外体运输。被子植物中既有导管又有管胞,裸子植物中只有管胞。导管是中空而无原生质体的长形死细胞,阻力小,运输速度较快,一般3~45m·h-1;而管胞中由于两管胞分子相连的细胞壁未打通,水分要经过纹孔才能在管胞间移动,所以阻力较大,运输速度比导管慢得多,不到0.6m·h-1。植物根部从土壤中吸收的水分进入根中柱之后,向地上部运输的途径是木质部的导管和管胞。它们占水分运输全部途径(从根表皮到叶表皮)的99.5%以上。水分在木质部中是以集流的方式沿着导管和管胞运输的。通常植物体内水分运输的速率是白天大于晚上,直射光下大于散射光下。
图 2-15 水分从根向地上部运输的途径
|
二、水分沿导管上升的机制
世界上高大的乔木如红杉、桉树可达100m以上,水分是靠什么力量从根部上升到最高叶片的呢?这涉及到两个问题,一是动力问题,二是水柱的连续性问题。
水分在导管中的运动是一种集流,其上升的动力为压力势梯度(即水势梯度),造成植株上下导管中压力势梯度的原因:一是根压(正压力势),二是蒸腾拉力(负压力势)。根压一般不超过0.2MPa, 至多能使水分上升20.4m。在一般情况下蒸腾拉力是水分上升的主要动力。强烈蒸腾时,顶端叶片水势可降至-3.0MPa,而根部水势一般在-0.2— -0.4MPa,因而根部的水分可顺着水势梯度上升至乔木的顶端。
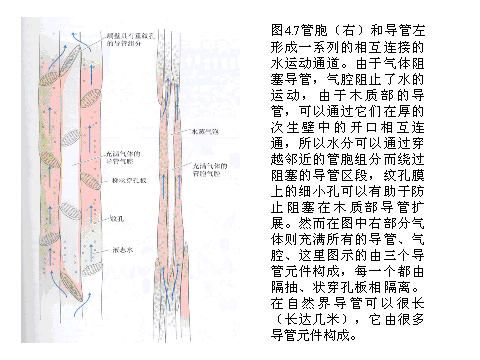
蒸腾作用产生的强大拉力把导管中的水往上拉,而导管中的水柱为何可以克服重力的影响而形成连续水柱而不中断呢?通常用爱尔兰人狄克逊(H.H.Dixon)提出的蒸腾流-内聚力-张力学说(transpirationcohesiontension theory)也称“内聚力学说”(cohesion theory)来解释的,即水分子的内聚力大于张力,从而能保证水分在植物体内的向上运输。水分子的内聚力很大,可达几十MPa。植物叶片蒸腾失水后,便向导管吸水,而水本身又有重量,会受到向下的重力影响,这样,一个上拉的力量和一个下拖的力量共同作用于导管水柱上就会产生张力(tension)。其张力可达-3.0MPa,但由于水分子内聚力远大于水柱张力,同时,水分子与导管(或管胞)壁的纤维素分子间还有强大的附着力,因而维持了输导组织中水柱的连续性,使得水分不断上升。但由于导管水溶液中溶解有气体,当水柱张力增大时,溶解的气体会从水中逸出而形成气泡。而且,在张力的作用下,气泡还会不断扩大。这种现象称为气穴现象(cavitation)。然而,植物可通过某些方式消除气穴造成的影响。例如当气泡在某些导管中形成后,它会被导管分子相连处的纹孔阻挡,气泡便被局限在一条管道中,当水分移动遇到了气泡的阻隔时,可以横向进入相邻的导管分子而绕过气泡,形成一条旁路,从而保持水柱的连续性。植物在夜间,由于蒸腾作用减弱,导管中水柱的张力也跟着降低,于是逸出的水蒸气或空气又可重新进入溶液,以解除气穴对水流的阻挡。另外,在导管内大水柱中断的情况下,水流仍可通过微孔以小水柱的形式上升。同时,水分上升也不需要全部木质部参与作用,只需部分木质部的输导组织畅通即可。
|
|
|
|
|
|