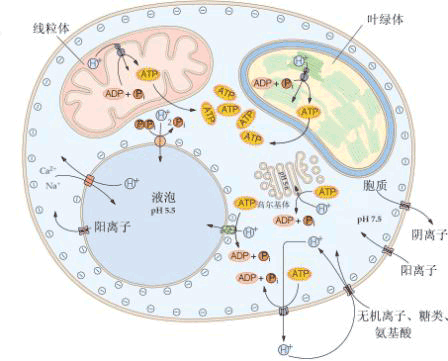
第二节 植物细胞对溶质的吸收
植物细胞可以从环境中吸收溶质,这个环境可以是植物生存的外部环境,如土壤,也可以是植物本身的内部环境,即一个细胞的周围组织。
当植物根内部的溶质浓度较低,从外部溶液吸收溶质时,首先有一个溶质迅速进入根的阶段,称为第一阶段,然后溶质吸收速度变慢且较平稳,这称为第二阶段(图3-2)。在第一阶段溶质通过扩散作用进入质外体,而在第二阶段溶质又进入原生质及液泡。如果将实验材料从溶液中取出再转入水中,则进入组织的溶质只有很小一部分会很快地泄漏出来,这就是原来进入质外体的部分。如果处于无O2、低温或用抑制剂来抑制呼吸作用,则第一阶段的吸收基本上不受影响,而第二阶段的吸收会被抑制(图3-2)。这些事实表明,溶质进入质外体与其跨膜进入细胞质和液泡的机制是不同的。前者是由于扩散作用而进行吸收,这种不需要代谢来提供能量的顺电化学势梯度吸收矿质的过程称为被动吸收(passive absorption)。后者要利用呼吸释放的能量才能逆电化学势梯度吸收矿质,这种过程称为主动吸收(active absorption)。
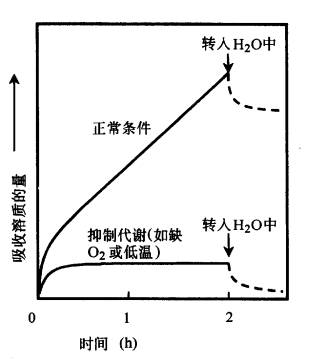 |
图3-2 植物组织对溶质的吸收
植物细胞吸收矿质元素的方式主要有二种类型:被动吸收和主动吸收。
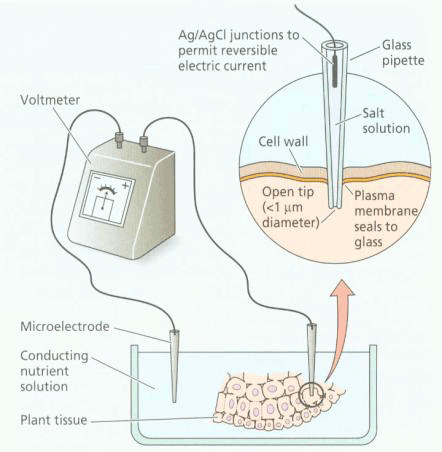
| 一、被动吸收 (一)扩散作用 扩散作用是指分子或离子沿着化学势或电化学势梯度转移的现象。电化学势梯度包括化学势梯度和电势梯度两方面,细胞内外的离子扩散决定于这两种梯度的大小,而分子的扩散则决定于化学势梯度或浓度梯度。 设细胞内的离子浓度为ai,膜电势为Ei,电化学势为μi;细胞外的离子浓度为a0,膜电势为Eo,电化学势为μo。根据电化学势差的表达式(2-10),细胞内外的电化学势差为
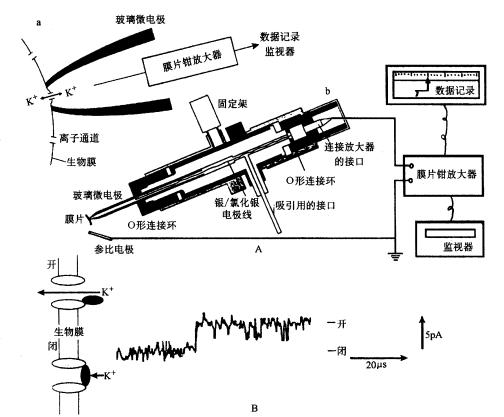
微电极插入细胞内,而面积大的参比电极安放在细胞外,二极间的电势差由静电计测量,电势差经放大器放大后由记录仪记录
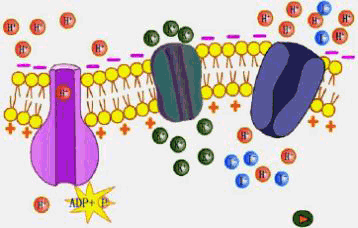
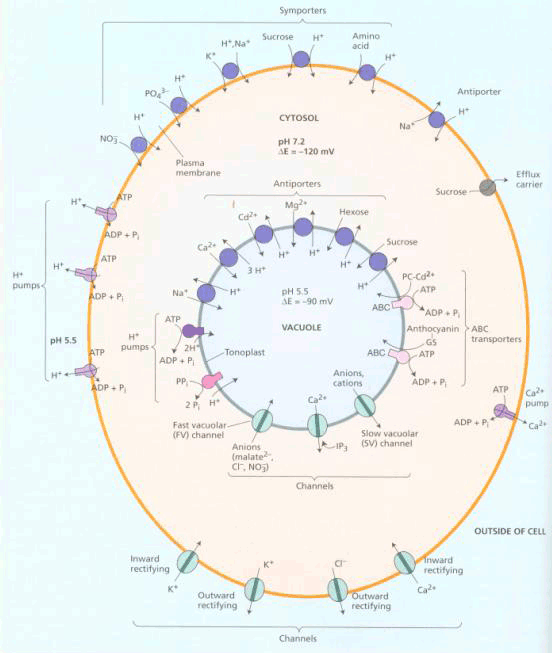
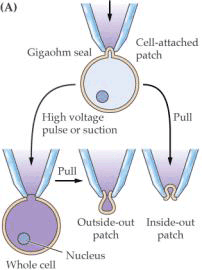
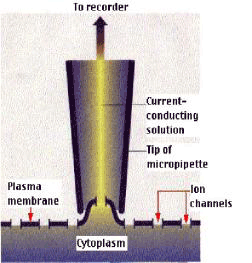
(二)协助扩散 协助扩散(facilitated diffusion)是小分子物质经膜转运蛋白顺浓度梯度或电化学梯度跨膜的转运。膜转运蛋白可分为两类:一类是通道(channel)蛋白,另一类是载体(carrier)蛋白。 1.离子通道(ion channel) 离子通道被认为是细胞膜中一类内在蛋白构成的孔道。可为化学方式或电学方式激活,控制离子通过细胞膜顺电化学势流动。 膜片钳(patch clamp,PC)技术的应用,极大地推动了对离子通道的研究。所谓膜片钳技术,是指使用微电极从一小片细胞膜上获取电子学信息的技术,即将跨膜电压保持恒定(电压钳位),测量通过膜的离子电流大小的技术。进行PC测定,需要用经过热抛光的尖端为1μm的玻璃微电极,压向清洁的膜表面,探截一小块膜,有时还需施以适当的吸力,目的是形成高阻封接。微电极中先灌适当的盐溶液,使通过细胞膜片的电流流入微电极。流入微电极的电流与探截膜片上的通道数目和开放状况以及通过的离子种类有关。微电极与高分辨力的放大器连接,根据记录到的电讯号,可推测离子通道的情况(图3-4)。膜片钳技术的试验材料往往是分离的原生质体或细胞器,这样可以避免细胞间的联系与多种细胞器的干扰,以便在较简单的环境中测定膜上通道特性。PC技术主要被用来分析膜上的离子通道,借此还可用来研究细胞器间的离子运输、气孔运动、光受体、激素受体以及信号分子等的作用机理,应用范围十分广泛。发明此技术的E.Neher和B.Sakmann荣获了1991年诺贝尔医学生理奖。
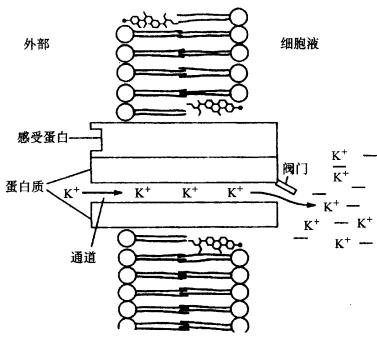
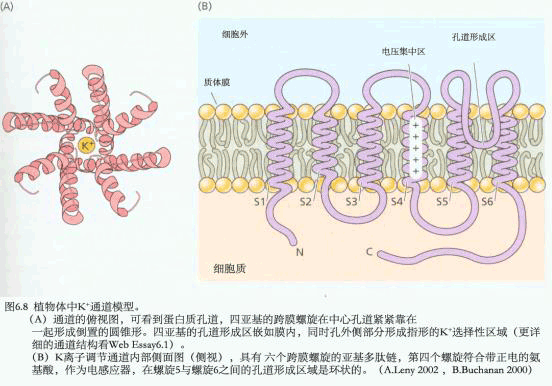
A.测定原理与装置:a.测定原理图,在玻璃微电极尖端截取的膜片上,如有开放的离子通道时,离子通过通道进入微电极,产生的电流经放大器放大后,由监视器显示或由记录仪记录;b.测定装置,示安装玻璃微电极的装置,有吸引接口和信号输出接口;B.通道开闭时的电流输出记录图,示仅通过一个离子通道时的膜电流情况,只有在通道开时才能测到电流。 现已观察到原生质膜中有K 、Cl-、Ca2 通道。原生质膜中也可能存在着供有机离子通过的通道。从保卫细胞中已鉴定出两种K 通道,一种是允许K 外流的通道,另一种则是吸收K 内流的通道,这两种通道都受膜电位控制。离子通道的构象会随环境条件的改变而发生变化,处于某些构象时,它的中间会形成孔,允许溶质通过。孔的大小及孔内表面电荷等性质决定了它转运溶质的选择性。根据孔开闭的机制可将通道分为两类:一类可对跨膜电势梯度发生反应,另一类则对外界刺激(如光照、激素等)发生反应。图3-5是一个假想的离子通道模型。在通道内进行的离子转运是顺着化学势或电化学势梯度的。跨膜的内部蛋白其中央孔道允许离子(K )通过。在这里, K 顺其电化学势梯度(注意通道右侧过量的负电荷)、但逆着浓度梯度从通道左侧(外)移向右侧(细胞质)。感受蛋白(sensor protein)可对细胞内外由光照、激素或Ca2 引起的化学刺激做出反应。通道上的阀门(gate)可以通过一种未知的方式对膜两侧的电势梯度或由环境刺激产生的化学物质做出开或关的反应。
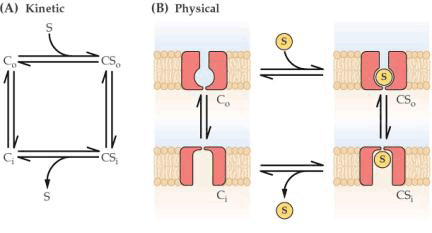
2.载体 载体也是一类内部蛋白,由载体转运的物质首先与载体蛋白的活性部位结合,结合后载体蛋白产生构象变化,将被转运物质暴露于膜的另一侧,并释放出去。由载体进行的转运可以是被动的(顺电化学势梯度),也可以是主动的(逆电化学势梯度),图3-6是一个通过载体进行被动转运的示意图。
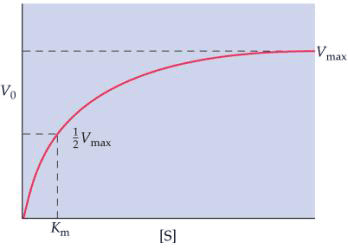
Km.载体与溶质的亲和力Vmax.最大速率 载体蛋白对被转运物质的结合及释放,与酶促反应中酶与底物的结合及对产物的释放情况相似。通过动力学分析,可以区别溶质是经通道还是经载体进行转运,经通道进行的转运是一种简单的扩散过程,没有饱和现象,而经载体进行的转运则依赖于溶质与载体特殊部位的结合,因结合部位的数量有限,所以载体转运有饱和现象(图3-7)。 |

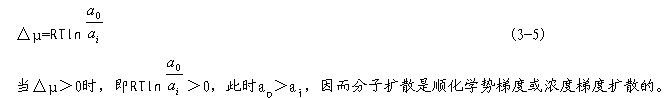
| 二、主动吸收 (一)ATP酶 通过载体的主动转运需要ATP提供能量。在高等植物根细胞质膜上存在着ATP酶(ATPase),它是1970年霍奇(Hodge)等用离体质膜小泡证实的。ATPase又称为ATP磷酸水解酶(ATP phosphorhydrolase),它可催化ATP水解生成ADP、磷酸,并释放能量,其反应如下:
A.B.ATP酶与细胞内的阳离子M 结合并被磷酸化; C.磷酸化导致酶的构象 改变,将离子暴露于外侧并释放出去; D.释放Pi恢复原构象 ATP酶是质膜上的插入蛋白(integral protein),它可以将ATP水解释放的能量用于转运离子。图3-8是一种ATP酶运送阳离子到膜外去的假设步骤。ATP酶上有一个与阳离子M 相结合的部位,还有一个与ATP的Pi结合的部位。当未与Pi结合时,M 的结合部位对M 有高亲合性,它在膜的内侧与M 结合,同时与ATP末端的Pi结合(称为磷酸化),并释放ADP。当磷酸化后,ATP酶处于高能态,其构象发生了变化,会将M 暴露于膜的外侧,同时对M 的亲合力降低,而将M 释放出去,并将结合的Pi水解释放回膜的内侧,这样ATP酶又恢复至原先的低能态构象,开始下一个循环。 由于这种转运造成了膜内外正、负电荷的不一致,所以形成了跨膜的电位差,故这种现象称为致电,又因为这种转运是逆电化学势梯度而进行的主动转运,所以也将ATP酶称为一种致电泵(electrogenic pump)。 不是所有的阳离子都以这种方式转运。H 是最主要的通过这种方式转运的离子,所以我们可以将转运H 的ATP酶称为H ATPase或H 泵。 质膜H ATPase的分子量100 000,其底物是MgATP,最适pH为6.5,最适温度30~40℃,K 能激活它的活性,比较专一的抑制剂为VO43-和己烯雌酚(DES)。 细胞质膜H ATPase是植物生命活动过程中的主宰酶(master enzyme),它对植物许多生命活动起着重要的调控作用,其在矿质元素转运中的主要作用是:①使细胞质的pH值升高,但由于细胞质有较强的缓冲作用,所以这种升高并不显著。通常细胞质的pH在7.0到7.5之间;②使细胞壁的pH值降低。由于胞壁的缓冲能力较小,其pH值通常降到5.0~5.5;③使细胞质相对于细胞壁表现电负性(electronegative)。这是由于将阳离子运出细胞质而保留阴离子,从而使质膜从内到外形成负的电势差的缘故。 液泡膜上也存在H ATP酶,其催化部分在细胞质一侧。在水解ATP过程中,它将H 泵入液泡,使液泡的电势通常比细胞质高20~30mV,而pH则为5.5。一些植物液泡的pH值可以更低,如柠檬(主要是液泡液)的pH为2.5,而海中的褐藻(Desmeretia)液泡的pH值可低于1.0。 液泡膜H -ATPase与质膜H -ATPase的区别:①其转运H 时不与ATP末端Pi结合 ②每水解一分子ATP运送两个H 进入液泡 ③不依赖于K 的激活,可被Cl-刺激 ④对钒酸盐不敏感,被NO-3抑制。此外叶绿体和线粒体膜上也存在有ATP酶。Ca2 是另一种通过ATP酶转运的离子。质外体中通常含较高浓度的Ca2 ,而细胞质中Ca2 浓度则十分低(微摩数量级)。Ca2 ATPase逆电化势梯度将Ca2 从细胞质转运到胞壁或液泡中。细胞质中Ca2 浓度很小的波动就会显著影响许多酶的活性,植物细胞可以通过调节Ca2 -ATPase的活性使细胞质中Ca2 保持一定水平。Ca2 ATPase的底物为Ca2 ATP,最适pH在7.0~7.5之间,受CaM等多种因素的调节。Ca2 ATPase可能只转运Ca2 。
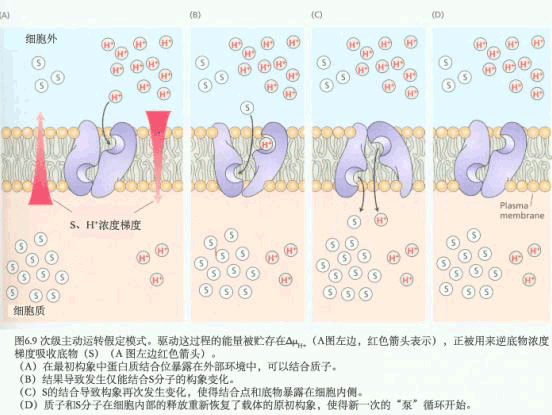
在线粒体与叶绿体中,用H 梯度中的能量来合成ATP,通过水解ATP与PPi的泵来建立跨膜的质子梯度。有这些泵建立的化学势被用来运输许多离子与小的代谢物穿过完整的膜通道与载体。 (二)共转运 质膜ATPase利用ATP水解产生的能量,把细胞质内的H 向膜外“泵”出。当质膜外介质中H 增加的同时,也产生膜电位(ΔE),即膜内呈负电性,而膜外呈正电性。跨膜的H 梯度和膜电位具有的能量合称为H 电化学势差,以 表示。根据(3-1)式, =RTln 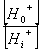 ZF(E0-Ei),因为H 的Z=1,E0-Ei=ΔE,因此;ΔμH =RTln ZF(E0-Ei),因为H 的Z=1,E0-Ei=ΔE,因此;ΔμH =RTln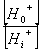 FΔE (3-6) FΔE (3-6)其中RTln 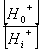 为化学势项,受H 浓度梯度影响。FΔE为电势项,受膜电位影响。ΔμH 也是其它种离子或分子越膜进入细胞以及由ATPase合成ATP(见第四章光合磷酸化)的驱动力。 为化学势项,受H 浓度梯度影响。FΔE为电势项,受膜电位影响。ΔμH 也是其它种离子或分子越膜进入细胞以及由ATPase合成ATP(见第四章光合磷酸化)的驱动力。
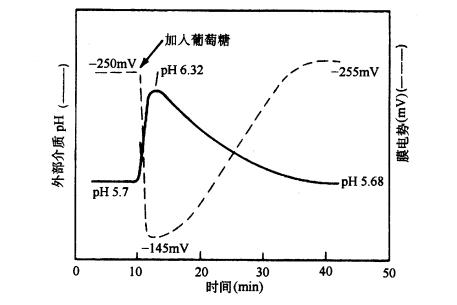
下面介绍一个葡萄糖/质子共向转运的实验证据(图3-10)。在简单的矿质营养液中培养水生植物膨胀浮萍(Lemno gibba),同时测定植物表面细胞的膜电势和培养液的pH。当向培养液中加入50mmol·L-1葡萄糖后,膜电势梯度的降低、外界培养液pH的升高与细胞吸收葡萄糖会同时发生。膜电势梯度的降低及外液pH的升高表明H 进入了细胞。但由于膜电势梯度的降低,会使H -ATP酶加速运转,从而又将更多H 运出细胞。这样经过一定时间后,膜电势梯度和外液pH又都恢复到原来水平。
(引自 Novacky,et al,1980 |