
植物呼吸作用多条途径都具有自动调节和控制能力。细胞内呼吸代谢的调节机理主要是反馈调节。所谓反馈调节(feedback regulation)就是指反应体系中的某些中间产物或终产物对其前面某一步反应速度的影响。凡是能加速反应的称为正效应物(positive effector)(正反馈物);凡是能使反应速度减慢者称负效应物(negative effector)(负反馈物)。对于呼吸代谢来说反馈调节主要是效应物对酶的调控,包括酶的形成(基因的表达)和酶的活性这两方面的调控,这里着重介绍反馈调节酶活性方面的内容(图5-18)。
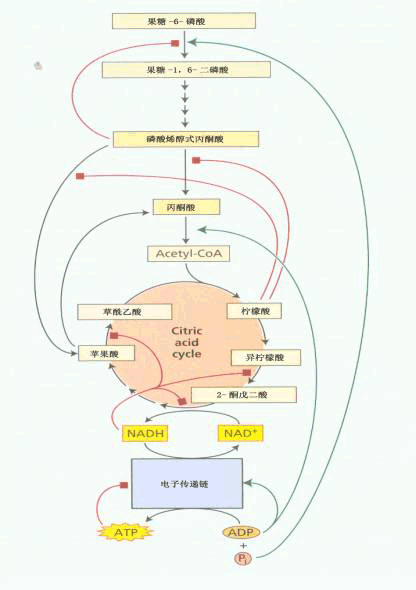 |
图5-18 呼吸代谢调节的可能部位
| 一、巴斯德效应和糖酵解的调节 当植物组织周围的氧浓度增加时,酒精发酵产物的积累逐渐减少,这种氧抑制酒精发酵的现象叫做“巴斯德效应”(Pasteur effect)。有氧条件下使发酵作用受到抑制是因为NADH的缺乏。在无氧条件下当3-磷酸甘油醛氧化为1,3-二磷酸甘油酸时,NAD 被还原成NADH H ;而当丙酮酸被还原为乳酸,乙醛被还原为乙醇时,NADH又被氧化成NAD ,如此循环周转。但在有氧条件下则不同,NADH能够通过GP—DHAP穿梭透入线粒体,用于呼吸链电子传递,因此NADH不能用于丙酮酸的还原,发酵作用就会停止。在有氧条件下糖酵解的速度减慢的原因是调节糖酵解的两个变构调节酶—磷酸果糖激酶和丙酮酸激酶在有氧条件下受到抑制的缘故。因为在有氧条件下,丙酮酸通过丙酮酸脱氢酶形成乙酰CoA,进入TCA循环,这样就会产生较多的ATP和柠檬酸,它们作为负效应物对两个关键酶起反馈抑制作用,糖酵解的速度自然就减慢了。这样就可以减少底物的消耗,把呼吸作用的速度自动控制在恰当的水平上。作为糖酵解两个关键酶的正效应剂有ADP、Pi、F1,6BP、Mg2 和K ,负效应剂还有Ca2 、3-磷酸甘油酸、2-磷酸甘油酸、磷酸烯醇式丙酮酸等。在无氧条件下,丙酮酸的有氧降解受到抑制,柠檬酸和ATP合成减少,积累较多的ADP和Pi,促进了两个关键酶活性,使糖酵解速度加快。此外,己糖激酶也参与调节糖酵解速度,属于变构调节酶,其变构抑制剂为其产物6-磷酸葡萄糖。 |
| 二、丙酮酸有氧分解的调节 丙酮酸在有氧条件下继续氧化的过程中,多种酶促反应受到反馈调节。首先是丙酮酸氧化脱羧酶系的催化活性受到乙酰CoA和NADH的抑制。这种抑制效应可相应地为CoA和NAD 所逆转。 TCA循环也受到许多因素的调节。过高浓度的NADH,对异柠檬酸脱氢酶、苹果酸脱氢酶等的活性均有抑制作用。NAD 为上述酶的变构激活剂。ATP对异柠檬酸脱氢酶、 α-酮戊二酸脱氢酶和苹果酸脱氢酶均有抑制作用,而ADP对这些酶有促进作用。琥珀酰CoA对柠檬酸合成酶和α-酮戊二酸脱氢酶有抑制作用。AMP对α-酮戊二酸脱氢酶活性,CoA对苹果酸酶活性都有促进作用。α-酮戊二酸对异柠檬酸脱氢酶的抑制和草酰乙酸对苹果酸脱氢酶的抑制则属于终点产物的反馈调节。此外,柠檬酸的含量可调节丙酮酸进入TCA循环的速度,柠檬酸多时,可以反馈抑制丙酮酸激酶,减少柠檬酸的合成。 |
| 三、PPP的调节 PPP主要受NADPH/NADP 比值的调节,NADPH竞争性地抑制葡萄糖-6-磷酸脱氢酶的活性,使葡萄糖-6-磷酸转化为6-磷酸葡萄糖酸的速率降低。NADPH也抑制6-磷酸葡萄糖酸脱氢酶活性。葡萄糖-6-磷酸脱氢酶也被氧化的谷胱甘肽所抑制。而光照和供氧都可提高NADP 的生成,可以促进PPP。植物受旱、受伤、衰老、种子成熟过程中PPP都明显加强,在总呼吸中所占比例加大。 |
| 四、能荷的调节 由ATP、ADP、AMP组成的腺苷酸系统是细胞内最重要的能量转换与调节系统。阿特金森(Atkinson,1968)提出“能荷”(energy charge,EC)的概念,它所代表的是细胞中腺苷酸系统的能量状态。细胞中由ATP、ADP和AMP三种腺苷酸组成的腺苷酸库是相对稳定的,它们易在腺苷酸激酶(adenylate kinase)催化下进行可逆的转变。通过细胞内腺苷酸之间的转化对呼吸代谢的调节作用称为能荷调节。可用下列方程式表示: 从上式可以看出,当细胞中全部腺苷酸都是ATP时,能荷为1;全部是AMP时,能荷为0,全部是ADP时,能荷为0.5。三者并存时,则能荷随三者比例的不同而异。 通过细胞反馈控制,活细胞的能荷一般稳定在0.75~0.95。反馈控制的机理如下:合成ATP的反应受ADP的促进和ATP的抑制;而利用ATP的反应则受到ATP的促进和ADP的抑制。如果在一个组织中需能过程加强时,便会大量消耗ATP,ADP增多,氧化磷酸化作用加强,呼吸速率增高,因而便大量产生ATP。相反,当需能降低时,ATP积累,ADP处于低水平,氧化磷酸化作用减弱,呼吸速率就下降。因而,细胞内的能荷水平可以调节植物呼吸代谢的全过程。 |
| 五、电子传递途径的调控 线粒体中电子传递途径会由于内外因的影响而发生改变。如处于稳定生长期的酵母细胞内线粒体在氧化NADH时,P/O是3;而处于稳定生长期前的P/O则是2,这说明二者的电子传递途径是不同的。大量实验证明,植物在感病、受旱、衰老时交替途径都有明显加强。马铃薯块茎的伤呼吸,刚开始的时候,切片呼吸的80%~100%是对CO及CN-敏感的,24h以后CO对切片的呼吸只起极小的作用,CN-的作用也减小。这表明,电子传递途径已由以细胞色素氧化系统为主的途径改变为对CN-和CO不敏感的抗氰途径。在植物体内,内源激素乙烯和内源水杨酸(salicylic acid)可诱导交替途径的运行,外源水杨酸和乙烯也能诱导交替途径的增强,同时可以诱导交替氧化酶基因的提前表达。植物缺磷时,体内ADP和Pi含量降低,磷酸化作用受到抑制,底物脱下的电子就越过复合体Ⅰ而直接交给UQ,并进入交替途径,以适应缺磷环境。 |
