| 第五节 植物细胞的基因表达 一、细胞的阶段性与全能性 繁殖、分化和衰亡是细胞的基本生命活动,也是细胞生理研究的重要内容,高等植物因细胞的这些基本生命活动而完成个体的生活史。 (一)细胞周期与细胞的阶段性 细胞繁殖(cell reproduction)是通过细胞分裂来实现的。从一次细胞分裂结束形成子细胞到下一次分裂结束形成新的子细胞所经历的时期称细胞周期(cell cycle),细胞周期所需的时间叫周期时间(time of cycle),整个细胞周期可分为间期(interphase)和分裂期两个阶段。间期是从一次细胞分裂结束到下一次分裂开始之间的间隔期。间期是细胞的生长阶段,其体积逐渐增大,细胞内进行着旺盛的生理生化活动,并作好下一次分裂的物质和能量准备,主要是DNA复制、RNA的合成与有关酶的合成以及ATP的生成。细胞周期可分为以下四个时期。 1.G1期 从有丝分裂完成到DNA复制之前的这段间隙时间叫G1期(gap1,pre-synthetic phase)。在这段时期中有各种复杂大分子包括mRNA、tRNA、rRNA和蛋白质的合成。 2.S期 这是DNA复制时期,故称S期(synthetic phase),这期间DNA的含量增加一倍。 3.G2期 从DNA复制完成到有丝分裂开始的一段间隙称G2期(gap2,post-synthetic phase),此期的持续时间短,DNA的含量不再增加,仅合成少量蛋白质。 4.M期 从细胞分裂开始到结束,也就是从染色体的凝缩、分离并平均分配到两个子细胞为止的时期。分裂后细胞内DNA减半,这个时期称M期(即有丝分裂,mitosis)或D期(division)。细胞分裂的意义在于S期中倍增的DNA以染色体形式平均分配到两个子细胞中,使每个子细胞都得到一整套和母细胞完全相同的遗传信息。 细胞周期延续时间的长短随细胞种类而异,也受环境条件的影响(见第八章第二节)。多细胞生物体要维持正常的生活,就必须不断地增殖新细胞以代换那些衰老死亡的细胞。 细胞衰老(cellular aging,见第十章第五节)与死亡是细胞生命活动的必然规律。细胞的死亡可以分为两种形式:一种是坏死性或意外性死亡(necrosis,accidental death),即细胞受到外界刺激,被动结束生命;另一种死亡方式称为程序化死亡(programmed cell death,PCD),这是一种主动的,受细胞自身基因调控的过程,因此,PCD是生命活动中不可缺少的组成部分。在PCD发生过程中,一般伴随有特定的形态、生化特征出现,此类细胞死亡被称为凋亡(apoptosis)。当然,也有的细胞在PCD过程中并不表现凋亡的特征,这一类PCD被称为非凋亡的程序化细胞死亡(non-apoptotic programmed cell death)。 细胞程序化死亡与通常意义上的细胞衰老死亡不同,它是多细胞生物中某些细胞所采取的一种自身基因调控的主动死亡方式。它与细胞坏死的形态特征也截然不同,其最明显的特征是细胞核和染色质浓缩,DNA降解成寡聚核苷酸片断,细胞质也浓缩,细胞膜形成膜泡,最后转化成凋亡小体(apoptotic body)。植物细胞程序化死亡主要发生在细胞分化过程中,如维管系统的发育;性别发生过程中某些生理器官的败育,导致单性花的形成。在发生过敏反应(hypersensitive reaction)时,细胞程序化死亡可在感染区域及其周围形成病斑(lesion),从而防止病原体的扩散。 (二)细胞分化与细胞全能性 细胞分化(cell differentiation)是细胞间产生稳定差异的过程。 也就是由一种类型的细胞转变成在形态结构、生理功能和生物化学特性诸方面不同的另一类型细胞的过程。植物体的各个器官以及各种组织内的细胞形态结构、功能和生理生化特性都是各不相同的,这就是细胞分化的结果。多细胞生物体的所有不同类型的细胞都是由受精卵发育而成的,正是在细胞分裂的基础上有了分化,才能使不同类型的细胞执行千差万别的生理代谢,共同完成植物的生命活动。 通过对细胞分裂过程的研究知道,母细胞在分裂前已经对DNA进行了复制,因而子细胞具有母细胞的全套基因。而植物体的所有细胞追踪溯源都来自受精卵,具有与受精卵相同的基因。细胞全能性(totipotency)就是指每个生活的细胞中都包含有产生一个完整机体的全套基因,在适宜的条件下,细胞具有形成一个新的个体的潜在能力。受精卵是全能的,它可以分裂繁殖和分化成各类细胞,并且能复制出一个完整植株。其他器官和组织的植物细胞,由于分裂和分化的结果,只具有其所在组织器官的特定功能,若要让其表现出全能性,就要了解其基因表达及调控机制。 |
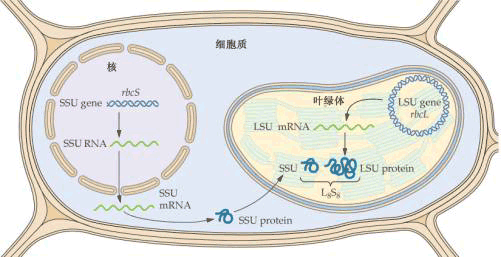
| 二、植物细胞的核基因与核外基因 高等植物细胞共有三个基因组,即核基因组、叶绿体基因组和线粒体基因组,后两组称为核外基因。基因组(genomes)是细胞携带生命信息DNA及其蛋白质复合物的总称。 以往普遍认为DNA只存在于细胞核中。1962年Ris和Plant在衣藻叶绿体中发现DNA。1963年M.Nass和S.Nass在鸡胚肝细胞线粒体中发现DNA,以后从植物细胞线粒体中也发现了DNA。 进一步研究发现,线粒体和叶绿体中还有RNA(mRNA、tRNA、rRNA)、核糖体、氨基酸活化酶等。说明这两种细胞器均有自我繁殖所必需的基本组分,具有转录和翻译的功能。然而,线粒体和叶绿体中的绝大多数蛋白质是由核基因编码,在细胞质核糖体上合成的。其本身编码合成的蛋白质并不多。也就是说,线粒体和叶绿体的自主程度是有限的,不完全的。因此,线粒体和叶绿体的生长和增殖是受核基因组及其自身的基因组两套遗传系统所控制的,所以称它们为半自主性细胞器(semiautonomous organelle)。 线粒体基因组也叫线粒体DNA(mtDNA),呈双链环状,与细菌DNA相似。一个线粒体中可有1个或几个DNA分子。酵母mtDNA的周长为26μm,有78 000个碱基对,高等植物mtDNA的大小在200000碱基对(油菜)和2 500 000碱基对(西瓜)之间。某些植物的线粒体中还含有较小的线形或环形DNA分子。叶绿体基因组也叫叶绿体DNA(cpDNA),双链环状,其大小差异在120 000~217 000个碱基对。cpDNA周长一般为40~60μm,分子量约为3.8×107,叶绿体中DNA的含量大体在1×10-14g水平上,明显地比线粒体中DNA含量(1×10-16g)多。每个线粒体中约含6个mtDNA分子,每个叶绿体中约含12个cpDNA分子。 mtDNA和cpDNA均可自我复制,其复制也是以半保留方式进行的。用3H嘧啶核苷标记证明,mtDNA复制时间主要在细胞周期的S期及G2期,DNA先复制,随后线粒体分裂,cpDNA复制的时间在G1期。它们的复制仍受核的控制,复制所需的DNA聚合酶常由核DNA编码并在细胞质核糖体上合成。 对1000种以上光合陆生植物的研究发现,叶绿体基因组在大小、结构、基因含量和整个基因构建上都是很保守的,现已对烟草、地钱和水稻等植物叶绿体DNA进行了全部顺序的测定,已知其大约有123个基因。这些基因大致可分为三类。一类是与光合过程有关的基因,如编码光系统I作用中心的A1、A2蛋白,光系统Ⅱ作用中心的D1、D2蛋白,ATP合成酶的一些亚基等。第二类是叶绿体基因表达所需的基因,如编码核糖体RNA、DNA聚合酶、RNA聚合酶等。第三类为其他基因,如编码NADH脱氢酶的若干亚基等。 叶绿体中的大部分多肽是由核基因编码并在细胞质的核糖体上合成的。细胞质中所合成的叶绿体中多肽的前体几乎都带有一段含几十个氨基酸序列的转运肽(transit peptide),这些前体由转运肽引导进入叶绿体后,转运肽被蛋白酶切去,同时相应的多肽到达预定部位。叶绿体内的相当组分都要求叶绿体基因和核基因的共同作用(表1-4)。光照则可促进或调节叶绿体基因的表达。 核酮糖-1,5-二磷酸羧化酶/加氧酶(ribulose-1,5-bisphosphate carboxylase/oxygenase,Rubisco,RuBPCO)就是叶绿体基因和核基因共同作用的典型例证。该酶是一个双功能酶,它既可催化羧化反应,又可催化加氧反应,在植物光合过程中的CO2同化及光呼吸中起着重要作用。高等植物的Rubisco是由8个大亚基和8个小亚基所构成的,其催化活性要依靠大、小亚基的共同存在才能实现。Rubisco大亚基由叶绿体DNA编码,并在叶绿体的核糖体上翻译,而小亚基则由核DNA编码,在细胞质核糖体上合成。Rubisco全酶由细胞质中合成的小亚基前体和叶绿体中合成的大亚基前体经修饰后组装而成(图1-18)。Rubisco约占叶绿体可溶性蛋白的50%,因此它也是自然界中最丰富的蛋白质。
Ssu.Rubisco小亚基; Lsu.Rubisco大亚基 已经研究的植物线粒体基因约有20余种,包括如下几类:一类是编码RNA的基因,如rRNA(18S,26S和5S),tRNA,如地钱mtRNA有29个tRNA基因;另一类是编码核糖体小亚基蛋白(如S12,S13)的基因;还有一类是某些酶复合物亚基的基因,如细胞色素氧化酶复合物的亚基Ⅰ、Ⅱ和Ⅲ、ATP酶复合物的部分亚基、NADH去氢酶的亚基等。同样,线粒体基因也要与核基因共同作用才能完成线粒体组分的表达过程。 |
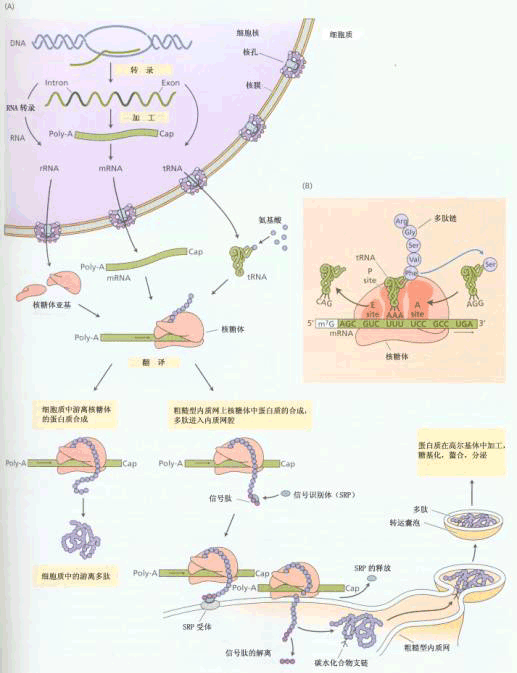
| 三、植物细胞基因表达的特点 基因的表达包括转录与翻译两个步骤。转录是RNA的生物合成,翻译是蛋白质的生物合成。 这两个过程都是很复杂的,且受到严格的调节控制。基因表达过程可用图1-19表示。
A.基因表达过程中的基本步骤,包括转录和翻译。蛋白质既可在游离于细胞质的核糖体上合成,又可在与膜结合的核糖体上合成。在细胞质基质中含有疏水的信号肽的分泌蛋白与信号识别体(signal recognition particle,SRP)结合,形成SRP核糖体复合体,后者转移到内质网上并与那里的SRP受体结合。随着翻译不断进行,延伸的多肽插入到内质网腔内,在那里信号肽解离,多肽与糖分子结合,形成的糖蛋白通过囊泡转运,被送到高尔基体,得到进一步的加工。 B.示tRNA携带氨基酸,以mRNA为模板,在核糖体 上延伸多肽链的翻译过程。(L. Taiz and E. Zeiger,Plant Physiology,2002) 基因表达调控在不同生物之间,尤其是原核生物与真核生物之间存在很大差异。这些差异主要归因于DNA在细胞中的存在形式、DNA分子的结构、参与基因表达的酶及蛋白因子等方面的差异。正是这些差异,决定了不同生物基因表达的各自特点。 (一)植物细胞中DNA的存在形式 原核细胞中的DNA是裸露的,遗传物质都集中在一条DNA分子上。而真核生物细胞的DNA含量和基因数目都远远多于原核细胞,其蛋白质或RNA的编码基因序列往往是不连续的,大多数基因都含有不表达序列,即内含子(intron)。另外,真核细胞DNA与组蛋白结合,以核小体为基本单位,形成念珠状长链,长链再高度压缩成为染色质或染色体。其遗传物质分散到多个DNA分子上。 在真核生物中,由于DNA与组蛋白等结合,形成核小体,阻碍了基因的转录,使DNA分子大部分区段常处于不表达的沉默态。因此,要使某一基因表达,必须去阻遏,打破沉默态。 (二)植物细胞DNA分子结构特点 原核生物细胞的DNA很少有间隔序列和重复序列,而真核细胞的DNA分子含有大量的重复序列。 原核生物的基因为多顺反子(polycistron),有操纵子(operon)结构。一些与功能相关的基因存在于同一操纵子中,受相同诱导物或阻遏物调节,这些基因的mRNA同时得以转录。真核生物的基因为单顺反子(cistron),无操纵子结构。基因有各自的调控序列,这些序列能够与调节蛋白结合,调控基因表达。 由于真核生物DNA结构上的复杂性和调节蛋白的多样性等原因,基因表达有明显的“时”与“空”的专一性。即有些基因,特别是与分化有关的基因,只在特定组织中表达,而且只在特定发育阶段表达。植物细胞基因表达的“时空”专一性主要表现在胚胎发生和花器发育两阶段,在这两个时期有基因的全新表达。 另外,与动物相比,植物基因表达更容易受环境因子(如光、温、水分)的影响,这些因子均可引起植物基因表达的改变。如以后要学习的植物的光周期现象、春化作用等都是环境因子作用于植物引起基因表达改变的明显例子。 (三)参与基因表达的酶及蛋白因子 原核生物中只有一种RNA聚合酶,该酶可以识别DNA分子上的转录起始部位,而真核生物中有三种RNA聚合酶。其中,RNA聚合酶I负责rRNA的合成,RNA聚合酶Ⅱ负责形成mRNA,该酶要在蛋白质(称为转录因子)参与下才能识别起始部位使转录开始。RNA聚合酶Ⅲ负责tRNA和小分子RNA的合成。 原核与真核生物中参与转录与翻译的蛋白因子也不同,且后者种类繁多。其中,真核生物的转录因子最具多样性。基因的时空专一性表达主要是转录因子识别基因的调控序列,并与RNA聚合酶相互作用,对内外环境因素作出反应的结果。 植物细胞中的DNA通过组蛋白阻遏(组蛋白经磷酸化等方式修饰)等机制,使大部分基因不能表达,又借在转录等水平上的各级复杂的调节机制,得以在特定组织和特定发育阶段中有相应基因进行适度表达,产生与组织结构和代谢功能相适应的蛋白和酶。这从理论上解释了植物细胞全能性在整株植物上被抑制,一般只能执行其所在组织的特定功能的原因。另一方面,从个体发育的观点上看,植物体每一细胞都来自受精卵,尤其是细胞有丝分裂的机制,保证了植物体的每一细胞都有相同的全套基因,这正是植物细胞具有全能性的内在遗传基础。 |