第三节 植物对矿质元素的吸收与运输
K 进入植株和在植株内运输的图解描绘 K 在木质部内的运输(红箭头)和在韧皮部中的运输(蓝箭头)。数字代表K 长距离运输途径中重要的运输位点。5个中的4个数字标示的位置,夸大描绘了K 在细胞水平的运输(1)K 被跨根细胞质膜吸收(纵切观察)。(2)K 通过跨木质部细胞膜输出。运输死的厚壁木质部导管(横切面观察)。 —、植物吸收矿质元素的特点 (一)根系吸收矿质与吸收水分不成比例 矿质元素必须溶解在水中,才能被植物吸收。过去认为植物吸收矿质是被水分带入植物体的。按照这种见解,水分和盐分进入植物体的数量,应该是成正比例的。但后来的大量研究证明,植物吸水和吸收盐分的数量会因植物和环境条件的不同而变化很大。有人用大麦作试验,通过光照来控制蒸腾,然后测定溶液中矿质元素的变化。结果发现,光下比暗中的蒸腾失水大2.5倍左右,但矿质吸收并不与水分吸收成比例(表3-6)。如磷酸根和钾离子在光下比暗中的吸收速度快,而其它无机盐,如Ca、Mg、SO2-4、NO-3等,在光下反而吸收少。总之,植物对水分和矿质的吸收是既相互关联,又相互独立。前者,表现为盐分一定要溶于水中,才能被根系吸收,并随水流进入根部的质外体。而矿质的吸收,降低了细胞的渗透勢,促进了植物的吸水。后者,表现在两者的吸收比例不同,吸收机理不同:水分吸收主要是以蒸腾作用引起的被动吸水为主,而矿质吸收则是以消耗代谢能的主动吸收为主。另外两者的分配方向不同,水分主要分配到叶片,而矿质主要分配到当时的生长中心。
(二)根系对离子吸收具有选择性 离子的选择吸收(selective absorption)是指植物对同一溶液中不同离子或同一盐的阳离子和阴离子吸收的比例不同的现象。例如供给NaNO3,植物对其阴离子(NO-3)的吸收大于阳离子(Na )。由于植物细胞内总的正负电荷数必须保持平衡,因此就必须有OH-或HCO3-排出细胞。植物在选择性吸收NO-3时,环境中会积累Na ,同时也积累了OH-或HCO-3,从而使介质pH值升高。故称这种盐类为生理碱性盐(physiologically alkaline salt),如多种硝酸盐。同理,如供给(NH4)2SO4,植物对其阳离子(NH 4)的吸收大于阴离子(SO2-4),根细胞会向外释放H ,因此在环境中积累SO2-4的同时,也大量地积累H ,使介质pH值下降,故称这种盐类为生理酸性盐(physiologically acid salt),如多种铵盐。如供给NH4NO3,则会因为根系吸收其阴、阳离子的量很相近,而不改变周围介质的pH,所以称其为生理中性盐(physiologically nutral salt)。生理酸性盐和生理碱性盐的概念是根据因植物的选择吸收引起外界溶液是变酸还是变碱而定义的。如果在土壤中长期施用某一种化学肥料,就可能引起土壤酸碱度的改变,从而破坏土壤结构,所以施化肥应注意肥料类型的合理搭配。 (三)根系吸收单盐会受毒害
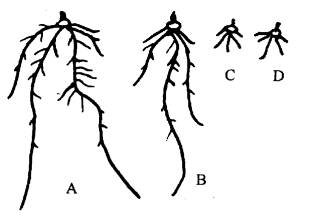
图 3-11小麦根在单盐溶液和盐类混合液中的生长状况 A.NaCl KCl CaCl;B.NaCl CaCl2; C.CaCl2; D.NaCl 任何植物,假若培养在某一单盐溶液中,不久即呈现不正常状态,最后死亡。这种现象称单盐毒害(toxicity of single salt)。单盐毒害无论是营养元素或非营养元素都可发生,而且在溶液浓度很稀时植物就会受害。例如把海水中生活的植物,放在与海水浓度相同的NaCl溶液中,植物会很快死亡。许多陆生植物的根系浸入Ca、Mg、Na、K等任何一种单盐溶液中,根系都会停止生长,且分生区的细胞壁粘液化,细胞破坏,最后变为一团无结构的细胞团。图3-11中的C和D显示了小麦根受单盐毒害的生长状况。 若在单盐溶液中加入少量其它盐类,这种毒害现象就会消除。这种离子间能够互相消除毒害的现象,称离子颉颃(ion antagonism),也称离子对抗或离子拮抗。所以,植物只有在含有适当比例的多盐溶液中才能良好生长,这种溶液称平衡溶液(balanced solution)。前边所介绍的几种培养液都是平衡溶液。对于海藻来说,海水就是平衡溶液。对于陆生植物而言,土壤溶液一般也是平衡溶液,但并非理想的平衡溶液,而施肥的目的就是使土壤中各种矿质元素达到平衡,以利于植物的正常生长发育。金属离子间的颉颃作用因离子而异,钠不能颉颃钾,钡不能颉颃钙,而钠和钾是可以颉颃钙和钡的。关于颉颃作用的本质,目前还没有满意的解释。 |

| 二、根系吸收矿质元素的区域和过程 (一)根系吸收矿质元素的区域
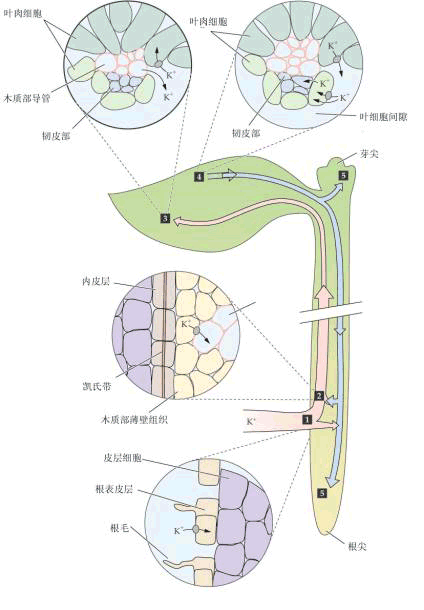
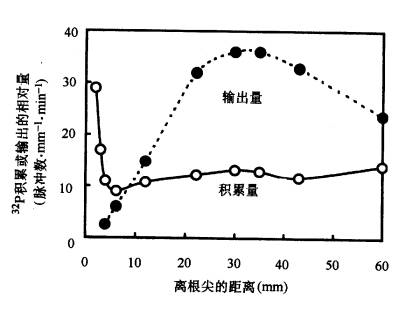
根系是植物吸收矿质的主要器官,它吸收矿质的部位和吸水的部位都是根尖未栓化的部分。过去不少人分析进入根尖的矿质元素,发现根尖分生区积累最多,由此以为根尖分生区是吸收矿质元素最活跃的部位。后来更细致的研究发现,根尖分生区大量积累离子是因为该区域无输导组织,离子不能很快运出而积累的结果;而实际上根毛区才是吸收矿质离子最快的区域,根毛区积累离子较少是由于离子能很快运出根毛区的缘故(图3-12)。 (二)根系吸收矿质的过程 根系吸收矿质要经过以下步骤: 1.离子被吸附在根系细胞的表面 根部细胞呼吸作用放出CO2和H2O。CO2溶于水生成H2CO3,H2CO3能解离出H 和HCO-3离子,这些离子可作为根系细胞的交换离子,同土壤溶液和土壤胶粒上吸附的离子进行离子交换,离子交换有两种方式: (1)根与土壤溶液的离子交换(ion exchange) 根呼吸产生的CO2溶于水后可形成的CO2-3、H 、HCO-3等离子,这些离子可以和根外土壤溶液中以及土壤胶粒上的一些离子如K 、Cl-等发生交换,结果土壤溶液中的离子或土壤胶粒上的离子被转移到根表面。如此往复,根系便可不断吸收矿质。如图3-13所示。 (2)接触交换(contact exchange) 当根系和土壤胶粒接触时,根系表面的离子可直接与土壤胶粒表面的离子交换,这就是接触交换。因为根系表面和土壤胶粒表面所吸附的离子,是在一定的吸引力范围内振荡着的,当两者间离子的振荡面部分重合时,便可相互交换。 离子交换按“同荷等价”的原理进行,即阳离子只同阳离子交换,阴离子只能同阴离子交换,而且价数必须相等。由于H 和HCO-3.分别与周围溶液和土壤胶粒的阳离子和阴离子迅速地进行交换,因此盐类离子就会被吸附在根表面。
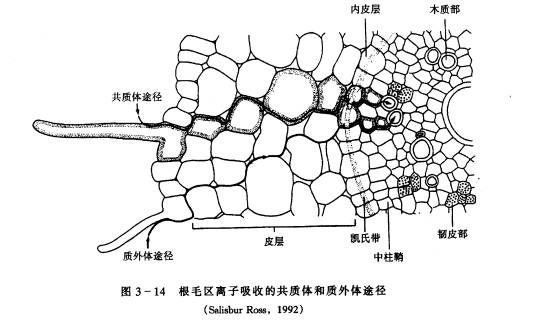
由于土壤颗粒表面带有负电荷,阳离子被土壤颗粒吸附于表面。外部阳离子如钾离子可取代土壤颗粒表面吸附的另一个阳离子如钙离子,使得Ca2 被根系吸收利用。 2.离子进入根部导管 离子从根表面进入根导管的途径有质外体和共质体两种(图3-14)。
图 3-14根毛区离子吸收的共质体和质外体途径 (Salisbur Ross,1992) (1)质外体途径 根部有一个与外界溶液保持扩散平衡、自由出入的外部区域称为质外体,又称自由空间。 自由空间的大小虽然无法直接测定,但可间接地推知其表观自由空间(apparent free space,AFS)的大小,即推知组织中自由空间的表观体积。测定时将根系放入某一物质的溶液中,待根内外离子达到平衡后,再测定溶液中的离子数和根内进入自由空间的离子数(将根再浸入水中,使自由空间内的离子扩散到水中,再行测定)。用下式可计算出表观自由空间:
各种离子通过扩散作用进入根部自由空间,但是因为内皮层细胞上有凯氏带,离子和水分都不能通过,因此自由空间运输只限于根的内皮层以外,而不能通过中柱鞘。离子和水只有转入共质体后才能进入维管束组织。不过根的幼嫩部分,其内皮层细胞尚未形成凯氏带前,离子和水分可经过质外体到达导管。另外在内皮层中有个别细胞(通道细胞)的胞壁不加厚,也可作为离子和水分的通道。 (2)共质体途径 离子通过自由空间到达原生质表面后,可通过主动吸收或被动吸收的方式进入原生质。在细胞内离子可以通过内质网及胞间连丝从表皮细胞进入木质部薄壁细胞,然后再从木质部薄壁细胞释放到导管中。释放的机理可以是被动的,也可以是主动的,并具有选择性。木质部薄壁细胞质膜上有ATP酶,推测这些薄壁细胞在分泌离子运向导管中起积极的作用。离子进入导管后,主要靠水的集流而运到地上器官,其动力为蒸腾拉力和根压。
胞间连丝如何连接相邻细胞中的细胞质的示意图 |


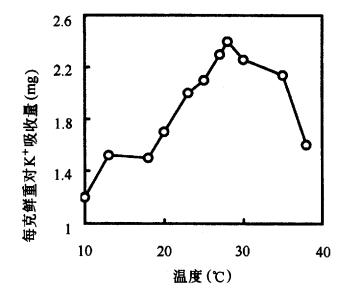
| 三、影响根系吸收矿质元素的因素 植物对矿质元素的吸收受环境条件的影响。其中以温度、氧气、土壤酸碱度和土壤溶液浓度的影响最为显著。 (一)温度 在一定范围内,根系吸收矿质元素的速度,随土温的升高而加快,当超过一定温度时,吸收速度反而下降(图3-15)。这是由于土温能通过影响根系呼吸而影响根对矿质元素的主动吸收。温度也影响到酶的活性,在适宜的温度下,各种代谢加强,需要矿质元素的量增加,根吸收也相应增多。原生质胶体状况也能影响根系对矿质元素的吸收,低温下原生质胶体粘性增加,透性降低,吸收减少;而在适宜温度下原生质粘性降低,透性增加,对离子的吸收加快。高温(40℃以上)可使根吸收矿质元素的速度下降,其原因可能是高温使酶钝化,从而影响根部代谢;高温还导致根尖木栓化加快,减少吸收面积;高温还能引起原生质透性增加,使被吸收的矿质元素渗漏到环境中去。
(二)通气状况 土壤通气状况直接影响到根系的呼吸作用,通气良好时根系吸收矿质元素速度快。根据离体根的试验,水稻在含氧量达3%时吸收钾的速度最快,而番茄必须达到5%~10%时,才能出现吸收高峰。若再增加氧浓度时,吸收速度不再增加。但缺氧时,根系的生命活动受影响,从而会降低对矿质的吸收。因此,增施有机肥料,改善土壤结构,加强中耕松土等改善土壤通气状况的措施能增强植物根系对矿质元素的吸收。土壤通气除增加氧气外,还有减少CO2的作用。CO2过多会抑制根系呼吸,影响根对矿质的吸收和其它生命活动。如南方的冷水田和烂泥田,地下水位高,土壤通气不良,影响了水稻根系的吸水和吸肥。 (三)土壤溶液浓度 据试验,当土壤溶液浓度很低时,根系吸收矿质元素的速度,随着浓度的增加而增加,但达到某一浓度时,再增加离子浓度,根系对离子的吸收速度不再增加。这一现象可用离子载体的饱和效应来说明。浓度过高,会引起水分的反渗透,导致“烧苗”。所以,向土壤中施用化肥过度,或叶面喷施化肥及农药的浓度过大,都会引起植物死亡,应当注意避免。 (四)土壤pH值 土壤pH值对矿质元素吸收的影响,因离子性质不同而异,一般阳离子的吸收速率随pH值升高而加速;而阴离子的吸收速率则随pH值增高而下降(图3-16)。
阴影面积的宽度表示植物体根系利用养分的程度。在PH5.5—6.5的范围内,所有的这些营养元素都可以被吸收利用。 vpH值对阴阳离子影响不同的原因,认为与组成细胞质的蛋白质为两性电解质有关,在酸性环境中,氨基酸带阳电荷,易吸收外界溶液中的阴离子;在碱性环境中,氨基酸带阴电荷,易吸收外部的阳离子。一般认为土壤溶液pH值对植物营养的间接影响比直接影响大得多。例如,当土壤的碱性逐渐增加时,Fe、Ca、Mg、Cu、Zn等元素逐渐变成不溶性化合物,植物吸收它们的量也逐渐减少;在酸性环境中,PO3-4、K 、Ca2 、Mg2 等溶解性增加,植物来不及吸收,便被雨水冲走。故在酸性红壤土中,常缺乏上述元素。另外,土壤酸性过强时,Al、Fe、Mn等溶解度增大,当其数量超过一定限度时,就可引起植物中毒。一般植物最适生长的pH值在6~7之间,但有些植物喜稍酸环境,如茶、马铃薯、烟草等,还有一些植物喜偏碱环境,如甘蔗和甜菜等。 |

| 四、叶片对矿质元素的吸收 植物除了根系以外,地上部分(茎叶)也能吸收矿质元素。生产上常把速效性肥料直接喷施在叶面上以供植物吸收,这种施肥方法称为根外施肥或叶面营养(foliar nutrition)。 溶于水中的营养物质喷施叶面以后,主要通过气孔,也可通过湿润的角质层进入叶内。角质层是多糖和角质(脂类化合物)的混合物,分布于表皮细胞的外侧壁上,不易透水。但角质层有裂缝,呈细微的孔道,可让溶液通过。溶液经过角质层孔道到达表皮细胞外侧壁后,进一步经过细胞壁中的外连丝到达表皮细胞的质膜。外连丝(ectodesmata)里充满表皮细胞原生质体的液体分泌物,从原生质体表面透过壁上的纤细孔道向外延伸,与质外体相接。当溶液经外连丝抵达质膜后,就被转运到细胞内部,最后到达叶脉韧皮部。外连丝是营养物质进入叶内的重要通道,它遍布于表皮细胞、保卫细胞和副卫细胞的外围。 营养物质进入叶片的量与叶片的内外因素有关。嫩叶比老叶的吸收速率和吸收量要大,这是由于二者的表层结构差异和生理活性不同的缘故。温度对营养物质进入叶片有直接影响,在30℃、20℃和10℃时,叶片吸收32P的相对速率分别为100、71和53。由于叶片只能吸收溶解在溶液中的营养物质,所以溶液在叶面上保留时间越长,被吸收的营养物质的量就越多。凡能影响液体蒸发的外界环境因素,如光照、风速、气温、大气湿度等都会影响叶片对营养物质的吸收。因此,向叶片喷营养液时应选择在凉爽、无风、大气湿度高的期间(例如阴天、傍晚)进行。
叶片微量氮素吸收过程简图, 根木质部转运分配的硝酸盐经硝酸转运器被叶肉细胞吸收到细胞质中,经硝酸还原酶作用还原为亚硝酸,亚硝酸和质子一起转运到细胞叶绿体中,在基质中亚硝酸还原酶还原作用转化为铵,铵经变谷氨酸合成酶的一系列作用转变为谷氨酸,谷氨酸再次进入细胞质。在天冬酰氨转移酶的作用下将氨基转移到天冬氨酸,最后,天冬酰氨合成酶将天冬酰酸转变为天冬酰胺,ATP值的大约数量就是每步反应上方所给的数值。 根外施肥具有肥料用量省、肥效快等特点,特别是在作物生长后期根系活力降低、吸肥能力衰退时;或因干旱土壤缺少有效水、土壤施肥难以发挥效益。或因某些矿质元素如铁在碱性土壤中有效性很低;Mo在酸性土壤中强烈被固定等情况下,采用根外追肥可以收到明显效果。常用于叶面喷施的肥料有尿素、磷酸二氢钾及微量元素。谷类作物生长后期喷施氮肥,可有效地增加种子蛋白质含量 根外施肥的不足之处是对角质层厚的叶片(如柑橘类)效果较差;喷施浓度稍高,易造成叶片伤害。 |
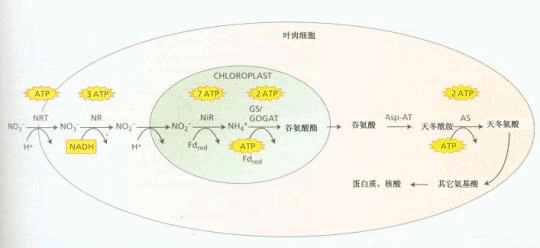
| 五、矿质元素在体内的运输和利用 吸收入根部的矿质元素,其中少部分留存在根内,大部分运输到植物的其它部位去,同样,被叶片吸收的矿质元素的去向与此雷同。
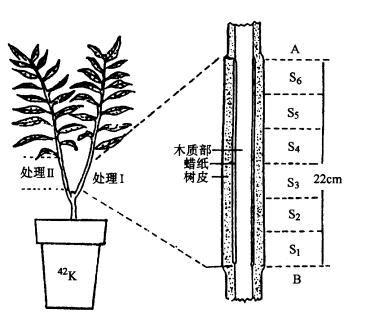
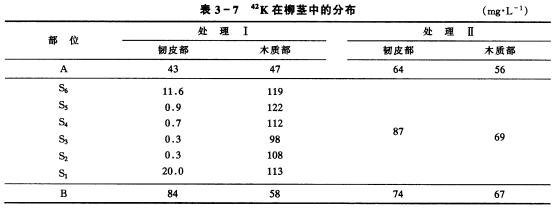
(一)矿质元素运输形式 根系吸收的N素,多在根部转化成有机化合物,如天冬氨酸、天冬酰胺、谷氨酸、谷氨酰胺,以及少量丙氨酸、缬氨酸和蛋氨酸,然后这些有机物再运往地上部;磷酸盐主要以无机离子形式运输,还有少量先合成磷酰胆碱和ATP、ADP、AMP、6磷酸葡萄糖、6磷酸果糖等有机化合物后再运往地上部;K 、Ca2 、Mg2 、Fe2 、SO2-4等则以离子形式运往地上部。 (二)矿质元素运输途径 将具有两个分枝的柳树苗,在两枝的对立部位把茎中的韧皮部和木质部分开(图3-17),在其中一枝的木质部与韧皮部之间插入蜡纸(处理Ⅰ),而另一枝不插腊纸,让韧皮部与木质部重新接触(处理Ⅱ),并以此作为对照。在根部施用42K,5小时后,再测定42K在茎中各部位的分布情况,结果如表3-7。可见在木质部内有大量42K,而在韧皮部内几乎没有,这表明根系所吸收的42K通过木质部的导管向上运输。在未分离区A与B处,以及分开后又重新将木质部与韧皮部密切接触的对照茎中,在韧皮部内也存在较多的42K显然这些42K是从木质部运到韧皮部的,由此表明矿质元素在木质部向上运输的同时,也可横向运输。 其它的试验证明,叶部吸收的矿质主要是通过韧皮部向下运输,也有横向运输。
矿质元素运到生长部位后,大部分与体内的同化物合成复杂的有机物质,如由氮合成氨基酸、蛋白质、核酸、磷脂、叶绿素等;由磷合成核苷酸、核酸、磷脂等;由硫合成含硫氨基酸、蛋白质、辅酶A等。它们进一步形成植物的结构物质。未形成有机化合物的矿质元素,有的作为酶的活化剂,如Mg、Mn、Zn等;有的作为渗透物质,调节植物水分的吸收。 已参加到生命活动中去的矿质元素,经过一个时期后也可分解并运到其它部位去,被重复利用。必需元素被重复利用的情况不同,N、P、K、Mg易重复利用,它们的缺乏病症,首先从下部老叶开始。Cu、Zn有一定程度的重复利用,S、Mn、Mo较难重复利用,Ca、Fe不能重复利用,它们的病症首先出现于幼嫩的茎尖和幼叶。氮、磷可多次参与重复利用;有的从衰老器官转到幼嫩器官(如从老叶转到上部幼叶幼芽);有的从衰老叶片转入休眠芽或根茎中,待来年再利用;有的从叶、茎、根转入种子中等等。矿质元素不只在植物体内从一个部位转移到另一个部位,同时还可排出体外。已知植物根系可以向土壤中排出矿质和其它物质;地上部分通过吐水和分泌也可将矿质和其它物质排出体外;另外下雨和结露能淋走植株中的许多物质,尤其是质外体中的物质。据报道,一年生植物在生长末期,钾的淋失可达最高含量的1/3,钙达1/5,镁达1/10;热带生长的籼稻在雨季淋失的氮可占所吸收氮的30%。可见,阴雨连绵会破坏植物体内的元素平衡。然而一些被淋洗和排出到土壤中的物质又可被根系再度吸收和利用。 |