无论是单细胞的藻类还是高大的树木,都存在体内同化物(assimilate)的运输和分配问题。叶片是同化物的主要制造器官,它合成的同化物不断地运至根、茎、芽、果实和种子中去,用于这些器官的生长发育和呼吸消耗,或者作为贮藏物质而积累下来。而贮藏器官中的同化物也会在一定时期被调运到其他器官,供生长所需要。同化物的运输与分配,无论对植物的生长发育,还是对农作物的产量和品质的形成都是十分重要的。
植物的生长发育除了受遗传基因控制外,还受到环境因素的影响,而且遗传基因的表达在很大程度上受到环境因素的调节。虽然植物有别于动物,不能逃避或改变环境,但植物经长期进化形成了一系列主动适应环境变化的机制,它可以感受外界环境信号,并通过某些机制将其转变为胞间和胞内信号,从而对细胞的代谢和基因的表达产生效应。因此,本章还将讨论植物细胞对环境信号的感受(perception)、植物体胞间及胞内信号传导(transduction)的有关内容。
高等植物体内的运输十分复杂,有短距离运输和长距离运输之分。短距离运输是指细胞内以及细胞间的运输,距离在微米与毫米之间;长距离运输是指器官之间、源库之间的运输,需要通过输导组织(有时称流),距离从几厘米到上百米。两者虽然都是物质在空间上的移动,但在运输的形式和机理上却有许多不同。这里说的源(source)即代谢源,指产生或提供同化物的器官或组织,如功能叶,萌发种子的子叶或胚乳;库(sink)即代谢库,指消耗或积累同化物的器官或组织,如根、茎、果实、种子等。应该指出,源库的概念是相对的,可变的,如幼叶是库,但当叶片长大长足时,它就成了源。 |
| 一、短距离运输系统 (一)胞内运输 胞内运输是指细胞内、细胞器间的物质交换。有分子扩散、微丝推动原生质的环流、细胞器膜内外的物质交换,以及囊泡的形成与囊泡内含物的释放等。如光呼吸途径中,磷酸乙醇酸、甘氨酸、丝氨酸、甘油酸分别进出叶绿体、过氧化体、线粒体;叶绿体中的丙糖磷酸经Pi转运器从叶绿体转移至细胞质,并在细胞质中合成蔗糖进入液泡贮藏;在内质网和高尔基体内合成的成壁物质由高尔基体分泌小泡运输至质膜,然后小泡内含物再释放至细胞壁中等这些过程均属胞内物质运输。 (二)胞间运输 胞间运输指细胞之间短距离的质外体、共质体以及质外体与共质体间的运输。 1.质外体运输 物质在质外体中的运输称为质外体运输(apoplastic transport)。质外体中液流的阻力小,物质在其中的运输快。由于质外体没有外围的保护,所以其中的物质容易流失到体外,另外运输速率也易受外力的影响。 2.共质体运输 物质在共质体中的运输称为共质体运输(symplastic transport)。由于共质体中原生质的粘度大,故运输的阻力大。在共质体中的物质有质膜的保护,不易流失于体外。 共质体运输受胞间连丝状态控制。一般认为,胞间连丝有三种状态: (1)正常态 内部具有固定的结构,能容许分子量小于1 000的小分子物质通过。 (2)开放态 连丝内部结构解体,扩大为开放的通道,足以让高分子通过。有一些病毒侵入细胞后,可诱发胞间连丝进入开放态,以利其自身能在细胞间转移,从而使感染区扩大。 (3)封闭态 连丝通道被粘液体等临时封闭,或永久堵塞,控制细胞内物质外运,并造成细胞间的生理隔离。
内吞作用:细胞外的物质通过吞噬(指内吞固体)或胞饮(指内吞液体)作用进入细胞质的过程;外排作用:将溶酶体或消化泡等囊泡内的物质释放到细胞外的过程; 出胞现象:通过出芽胞方式将胞内物质向外分泌的过程 一般地说,细胞间的胞间连丝多、孔径大,存在的浓度梯度大,则有利于共质体的运输。
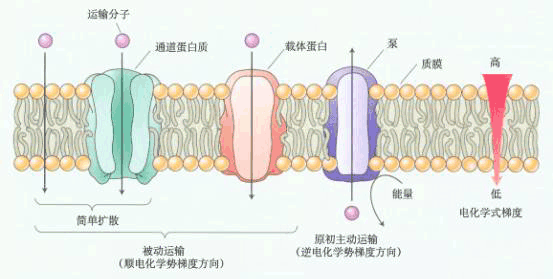
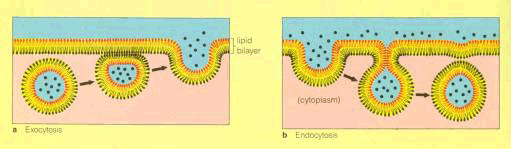
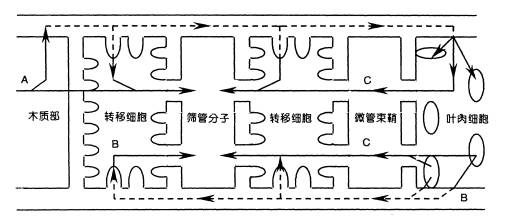
图 6-3 胞间运输途径示意图 实线箭头表示共质体途径,虚线箭头为质外体途径。A为蒸腾流,B为同化物在共质体质外体交替运输, C为共质体运输。 3.质外体与共质体间的运输 即为物质进出质膜的运输。物质进出质膜的方式有三种:(1)顺浓度梯度的被动转运(passive transport),包括自由扩散和通过通道或载体的协助扩散;(2)逆浓度梯度的主动转运(active transport)(图6-1),包括一种物质伴随另一种物质进出质膜的伴随运输;(3)以小囊泡方式进出质膜的膜动转运(cytosis),包括内吞(endocytosis)、外排(exocytosis)和出胞等(图6-2)。 植物体内物质的运输常不局限于某一途径。如共质体内的物质可有选择地穿过质膜而进入质外体运输;在质外体内的物质在适当的场所也可通过质膜重新进入共质体运输(图6-3)。这种物质在共质体与质外体之间交替进行的运输称共质体-质外体交替运输。 在共质体-质外体交替运输过程中常涉及一种特化细胞,起转运过渡作用,这种特化细胞被称为转移细胞(transfer cells,TC),它在结构上的特征是:细胞壁及质膜内突生长,形成许多折叠片层(见《植物学》)。这将扩大质膜的表面积,暴露更多的“溶质泵”或载体部位,从而增加溶质内外转运的面积;另外,质膜折叠能有效地促进囊泡的吞并,加速物质的分泌或吸收(电镜下观察到在转移细胞的质膜附近集中大量的囊泡)。 |
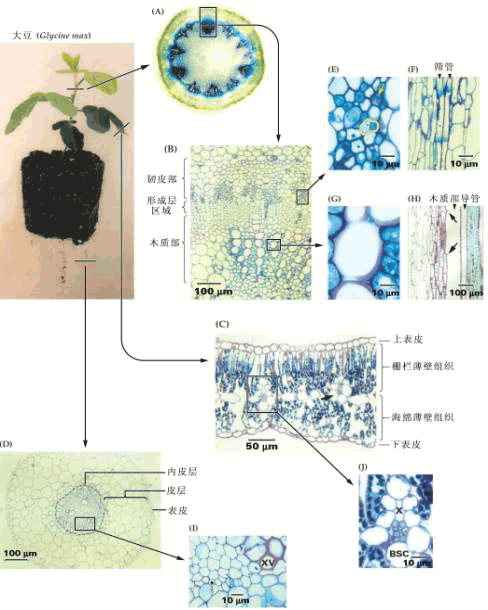
| 二、长距离运输系统 植物体内承担物质长距离运输的系统为维管束系统。 (一)维管束的组成与功能
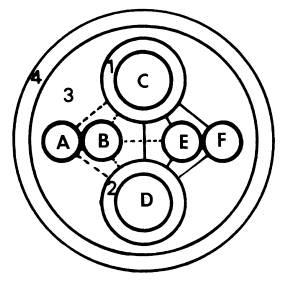
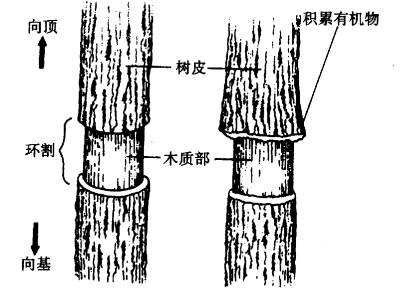
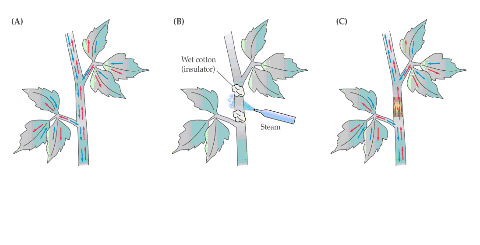
图 6-4 维管束的组成与功能 1.以导管为中心的木质部; 2.以筛管为中心的韧皮部; 3.多种组织的集合; 4.维管束鞘; A.电波; B.激素; C.无机营养; D.有机营养; E.加工储藏; F.径向生长; 实线表示物质交换,虚线表示信息交换 1.组成 一个典型的维管束可由四部分组成(图6-4):(1)以导管为中心,富有纤维组织的木质部。(2)以筛管为中心,周围有薄壁组织伴联的韧皮部。(3)穿插与包围木质部和韧皮部的多种细胞。(4)维管束鞘。 2.功能 通常认为维管束系统具有以下功能: (1)物质长距离运输的通道 一般情况下水和无机营养由木质部输送,而同化物则由韧皮部输送。 (2)信息物质传递的通道 如根部合成的细胞分裂素类和脱落酸等可通过木质部运至地上部分,而茎端合成的生长素则通过韧皮部向下极性运输。植物受环境刺激后产生的电波也主要在维管束中传播。 (3)两通道间的物质交换 木质部和韧皮部通过侧向运输可相互间运送水分和养分。如筛管中的膨压变化就是由于导管与筛管间发生水分交流引起的。 (4)对同化物的吸收和分泌 韧皮部对同化物的吸收与分泌,不仅发生在源库端,而且在同化物运输的途中维管束也能与周围组织发生物质交换。 (5)对同化物的加工和储存 同化物在运输过程中可卸至维管束中的某些薄壁细胞内,在其中合成淀粉,并储存下来。这些薄壁细胞就成为同化物的中间库,当其它库需要时,中间库的淀粉则可水解再转运出去。 (6)外源化学物质以及病毒等传播的通道 一些杀虫剂、灭菌剂、肥料以及病毒分子经两通道的传输,能产生周身效应。另外筛管汁液中还有一些蛋白抑制剂,可抑制动物消化道内的蛋白酶,这说明筛管本身存在一定的防卫机制。 (7)植物体的机械支撑 植物的长高加粗与维管束有密切关系,若树木没有木质部形成的心材,就不可能长至几米、几十米、甚至一百多米的高度。 当然在这些功能中,韧皮部最基本的功能是在源端把同化物装入筛管,在库端把同化物卸至生长细胞或贮藏细胞,以及提供同化物长距离运输的通道。 (二)物质运输的途径 1.研究物质运输途径的方法 木质部和韧皮部是进行长距离运输的两条途径,实验证明,同化物的运输是由韧皮部担任的。 (1)环割试验(girdling experiment) 这是研究物质运输的经典方法。环割是将树干(枝)上的一圈树皮(韧皮部)剥去而保留树干(木质部)的一种处理方法。此处理主要阻断了叶片形成的光合同化物经韧皮部向下运输,导致环割上端韧皮部组织中光合同化物积累引起膨大,环割下端的韧皮部组织因得不到光合同化物而死亡。如用水蒸气或冷却处理韧皮部组织均可观察到类似的情况。然而,必须指出的是,向下运输的韧皮部汁液中还含有许多含氮化合物、激素等,它们都能促进细胞的分裂和生长,与韧皮部组织膨大有关。如果环割不宽(如0.3~0.5cm),切口能重新愈合。如果环割较宽,环割下方又没有枝条,时间一久,根系就会死亡,这就是所谓的“树怕剥皮”。
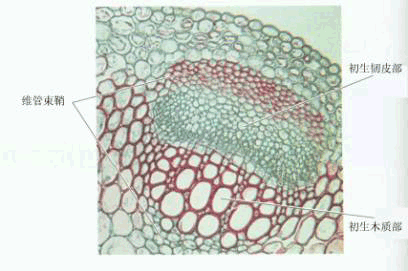
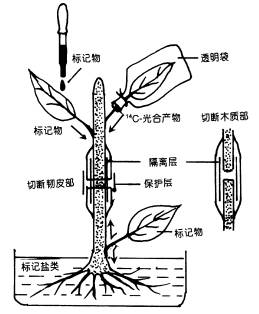
图6-5 韧皮部环割试验示意图 (2)同位素示踪法 采用放射性同位素示踪法,能更好地了解植物体内同化物运输的情况。如图6-6所示,可用几种方法将标记物质引入植物体。①根部标记32P、35S等盐类以便追踪根系吸收的无机盐类的运输途径 ②让叶片同化14CO2,可追踪光合同化物的运输方向 ③将标记的离子或有机物用注射器等器具直接引入特定部位。 标记一定时间后,将植株材料迅速冷冻、干燥(以防止标记物移动),用石蜡或树脂包埋,切成薄片,在薄片上涂一层感光乳胶,置于暗处,经过一段时间后,标记元素的辐射使乳胶片曝光,显定影后,胶片上与组织中存在标记元素的部位便会出现银颗粒(底片呈黑色处)。图6-7是对甜菜叶片进行14CO2标记, 10min后将叶柄切下并固定,对叶柄横切面进行放射性自显影的结果,从中可以看出14C标记的光合同化物位于韧皮部。若用14C标记的蔗糖直接饲喂除去表皮或蜡质层的叶片,也得到了同样的结果。采用上述同位素标记法还可测定物质运输速率,只要在运输的途径上(如茎杆上)相隔一定距离放置些检测放射性强度的探头,测定标记同位素经过相邻两个探头间的时间,便能推算运输速率。
图6-6 植物体内运输途径试验的示意图 将韧皮部和木质部剥离后插入蜡纸或胶片等不通透 的薄物制造屏障,以防止两通道间物质的侧向运输。 环割处理在实践中有多种应用。例如,对苹果、枣树等果树的旺长枝条进行适度环割,使环割上方枝条积累糖分,提高C/N比,促进花芽分化,提高坐果率,控制徒长。又如,在进行花木的高空压条繁殖时,可在欲生根的枝条上环割,在环割处附上湿土并用塑料纸包裹,由于此处理能使养分和生长素集中在切口上端,故利于发根。另外在用改良半叶法测定双子叶植物的光合速率时也需在测定叶的下方进行环割处理(对单子叶植物可进行化学环割,即用三氯乙酸等蛋白质沉淀剂涂叶枕下叶鞘以杀死韧皮细胞),防止叶中光合产物的外运。
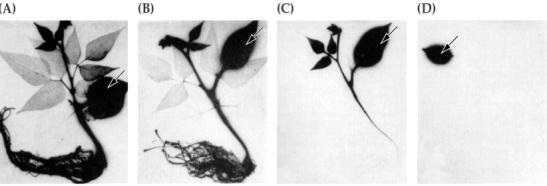
图 6-7 显示溶质在韧皮部中分布模式的全植株的放射自显影。 向同一大豆植株的幼叶(箭头)连续施加可在韧皮部中运动的放射性磷(如K2H32PO4)韧皮部,而其在24小时后会放射自显影。溶质向植物的生长部位运动,经过成熟的,蒸腾(光合反应)的叶片。(A和B中成熟叶片中的光标记可能代表从根中再循环的32P) 2.物质运输的一般规律 经大量研究,得到了有关物质运输途径与方向的一般性结论: (1)无机营养在木质部中向上运输,而在韧皮部中向下运输 (2)光合同化物在韧皮部中可向上或向下运输,其运输的方向取决于库的位置 (3)含氮有机物和激素在两管道中均可运输,其中根系合成的氨基酸、激素经木质部运输;而冠部合成的激素和含氮物则经韧皮部运输 (4)在春季树木展叶之前,糖类、氨基酸、激素等有机物可以沿木质部向上运输 (5)在组织与组织之间,包括木质部与韧皮部间,物质可以通过被动或主动转运等方式进行侧向运输 (6)也有例外的情况发生。 3.韧皮部运输 韧皮部是由筛管、伴胞和薄壁细胞组成的。虽然成熟的筛管分子也含有细胞质,但在分化过程中筛管分子细胞质中的细胞核、液泡、核糖体、高尔基体以及微管、微丝等细胞器被降解消失,而仅留下质膜、内质网、质体和线粒体等。筛管的细胞质中含有多种酶,如和糖酵解有关的酶,胼胝质合成酶,还含有韧皮蛋白(phloem protiein ,P-蛋白)和胼胝质等。 P-蛋白主要位于筛管的内壁。当韧皮部组织受到损伤时,处于高膨压状态下的筛管分子中的细胞质的正常状态就受到破坏,细胞内含物会迅速向受伤部位移动,这样P蛋白就会在筛孔周围累积并形成凝胶,堵塞筛孔以维持其他部位筛管的正压力,同时减少韧皮部内运输的同化物的不必要的损失。 胼胝质(callose)是一种以β1,3-键结合的葡聚糖。正常条件下,只有少量的胼胝质沉积在筛板的表面或筛孔的四周。但当植物受到外界刺激(如机械损伤、高温等)时,筛管分子内就会迅速合成胼胝质,并沉积到筛板的表面或筛孔内,堵塞筛孔,以维持其他部位筛管正常的物质运输。一旦外界刺激解除,沉积到筛板表面或筛孔内的胼胝质则会迅速消失,使筛管恢复运输功能。 筛管(sieve tube)是有机物运输的主要通道。由于筛管分子(sieve element,SE)间有筛板,而筛板的筛孔口径小,且常含有胼胝质,再加之筛管中的糖液浓度较大,因此,筛管流阻力较大,同化物在其中的流速较慢。 筛管通常与伴胞配对,组成筛管分子-伴胞复合体(sieve elementcompanion cell,SE-CC)。在源端或库端筛管周围不仅有伴胞,而且增加了许多薄壁细胞。茎中的伴胞比筛管分子小,而在源端或库端的伴胞或薄壁细胞的体积通常比筛管分子大,而且这些伴胞或薄壁细胞的细胞质浓,线粒体密度大,呼吸旺盛,代谢活跃,在功能上与茎中的伴胞不同,SE-CC在筛管吸收与分泌同化物以及推动筛管物质运输等方面起着重要的作用。 |