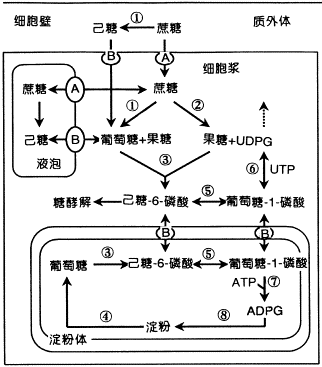
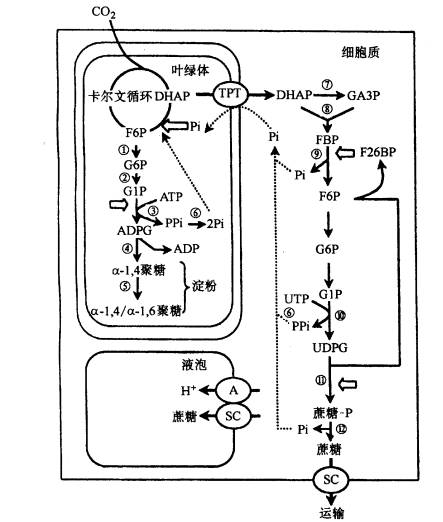
| 一、光合细胞中同化物的相互转化 淀粉和蔗糖是光合作用的主要终产物。光合碳代谢形成的磷酸丙糖的去向直接决定了有机碳的分配(图6-17)。磷酸丙糖或者继续参与卡尔文循环的运转;或者滞留在叶绿体内,并在一系列酶作用下合成淀粉;或者通过位于叶绿体被膜上的磷酸丙糖转运器(triose phosphate translocator,TPT)进入细胞质,再在一系列酶作用下合成蔗糖。合成的蔗糖或者临时贮藏于液泡内,或者输出光合细胞,经韧皮部装载通过长距离运输运向库细胞。因此,了解光合细胞中碳水化合物间的相互转化及其调节机理对阐明同化物的运输机理和调节具有重要的理论意义。
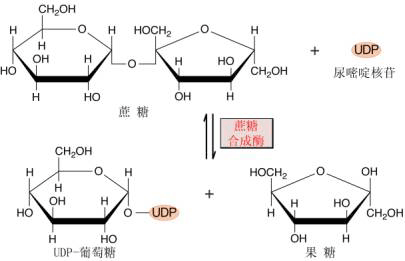
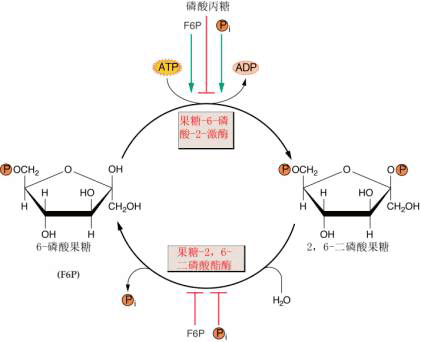
图6-17 光合细胞内淀粉和蔗糖的合成 (一)蔗糖的合成及调节 蔗糖是高等植物碳水化合物运输和贮藏的主要形式,也是协调植物源库关系的信号分子。因此蔗糖在植物生长发育和同化物分配中起着极其重要的作用。 1.合成部位和途径 光合细胞中参与蔗糖生物合成的所有酶位于细胞质内,据此认为,蔗糖的合成是在细胞质内进行的。光合中间产物磷酸丙糖通过叶绿体被膜上的磷酸丙糖转运器进入细胞质。在细胞质中,磷酸二羟丙酮(DHAP)在磷酸丙糖异构酶作用下转化为磷酸甘油醛(GAP),两者处于平衡状态,然后,DHAP和GAP在醛缩酶催化下形成果糖-1,6-二磷酸(F1,6BP)。F1,6BP C1位上的磷酸由果糖-1,6-二磷酸酯酶(FBPase)水解形成F6P。这一步反应是不可逆的,也是调节蔗糖合成的第一步反应。F6P在磷酸葡萄糖异构酶和磷酸葡萄糖变位酶作用下,形成G6P和G1P,此三种磷酸己糖处于动态平衡状态。然后由G1P和UTP合成蔗糖所需的葡萄糖供体UDPG和PPi, 反应是由UDPG焦磷酸化酶(UDP glucose pyrophosphory lase UGP)催化,这一步反应虽然是可逆的,但由于焦磷酸可被用于驱动位于液泡膜上的质子泵,以促使该反应向有利于蔗糖合成的方向进行(图6-17)。UDPG和F6P结合形成蔗糖-6-磷酸(S6P),催化该反应的酶是蔗糖磷酸合成酶(sucrose phosphate synthase SPS),它是蔗糖合成途径中另一个重要的调节酶。蔗糖合成的最后一步反应是S6P由蔗糖磷酸酯酶(sucrose phosphate phosphatase)水解形成蔗糖。 2.调节酶 蔗糖合成途径中至少有两步酶促反应对蔗糖的合成具有调节作用。 (1)果糖1,6-二磷酸酯酶(FBPase)和F2,6BP FBPase催化下列反应: F1,6BP+H2O→F6P Pi 该酶具有下列特性: ①对底物F1,6BP具有很高的亲和力,Km为2~4μmol·L-1; ②催化活性需Mg2 和中性pH; ③果糖-2,6-二磷酸(F2,6BP)不仅是该酶的强抑制剂,而且还改变了FBPase催化反应动力学特性。另外F2,6BP还能加强AMP对FBPase的抑制活性以及提高该酶对pH和Mg2 的依赖性。因此,普遍认为F2,6BP在调节FBPase和蔗糖合成中起关键性的作用。
图 6-18 6-磷酸果糖-2-激酶和果糖-2,6-二磷酸酶作用下的2,6-二磷酸果糖的形成和降解。 无机磷、丙糖磷酸和6-磷酸果糖起调节作用 细胞内F2,6BP的浓度决定于果糖-2,6-二磷酸酯酶(F2,6BPase)和果糖-6-磷酸-2-激酶(F6P,2K)的活性。 现已证明,F2,6Bpase和F6P,2K这两种酶位于细胞质中。Pi和F6P可提高F6P,2K的Vmax 和对F6P的亲和性,而三碳磷酸化中间产物如3PGA、磷酸丙糖、PEP则抑制该酶活性。催化F2,6BP水解的F2,6BPase则受Pi和F6P的抑制(图6-18)。 (2 蔗糖磷酸合成酶(SPS) SPS催化UDPG与F6P结合形成磷酸蔗糖: UDPG F6P蔗糖6P UDP H SPS具有以下特性: ①含量低和稳定性差。SPS在叶片中的含量只占总可溶性蛋白的0.1%以下,离体酶很不稳定,易失活 ②由2个亚单位组成,每个亚单位的分子量在120 000~138 000之间 ③催化反应所需的最适pH为中性 ④对UDPG和F6P的Km分别为3.0mmol.l-1和2.0mmol·L-1 ⑤SPS受G6P和Pi的异构调节,G6P提高V 和酶对F6P的亲和性,而Pi则抑制该酶的活性 ⑥SPS是一个光调节酶,其调节机理与SPS蛋白质158位丝氨酸的磷酸化(低活性)和去磷酸化(高活性)有关 ⑦SPS还受渗透调节。高渗溶液可提高该酶的活性。 (二)蔗糖合成对光合作用和同化物分配的调节 光合作用的正常进行不但要求足够量的ATP的供应,而且需要光合中间产物的数量维持一定的水平,这两者的数量与细胞质中Pi的水平密切相关。Pi太低,磷酸丙糖(TP)通过Pi转运器进入细胞质的量减少。在这种情况下,卡尔文循环中间产物的量虽然能维持较高水平,但由于叶绿体间质中Pi水平的下降导致ATP合成受阻,反过来抑制光合作用。Pi太高,磷酸丙糖输出的量超过了维持卡尔文循环正常运转所需的用于再生RuBP的量,从而又会引起光合碳代谢速率的下降。因此,Pi在叶绿体间质和细胞质之间有效的再循环,对维持光合作用的正常运行是必需的,这要求蔗糖和其他物质如淀粉、氨基酸的合成速率与光合作用的速度相互协调。 在光合细胞中,这种协调作用是通过FBPase和SPS的调节特性达到的。这里主要讨论两种情况:一种情况是当光合作用形成的TP速度超过蔗糖合成速度时,蔗糖的合成受前馈激活(feedforward activation); 另一种情况是蔗糖合成速度超过光合速率时,蔗糖的合成受反馈抑制(feedback inhibition)。 当光合作用形成TP的速度超过蔗糖合成速度时,TP通过Pi转运器进入细胞质的速率增加,降低了细胞质中Pi的水平。低Pi和高ATP降低了F6P,2K的活性,也抑制了F2,6Pase的活性,从而F2,6P合成速率下降,而降解速率加快,导致细胞质中F2,6BP水平降低。同时,F6P和G6P合成速率增加。G6P浓度的增加和Pi浓度的降低直接提高了SPS的活性,抑制了SPS激酶的活性,解除了对蔗糖磷酸磷酸酯酶的抑制,最终蔗糖合成速率增加直至蔗糖合成速率与光合速率达到新的平衡。这种由于光合速率提高促进蔗糖合成的现象称前馈激活。当光合速率降低或蔗糖合成速率过高时,则情况正好相反,细胞质蔗糖的合成受到反馈抑制,直至蔗糖合成速率与光合速率相协调为止(图6-18)。 (三)光合细胞内蔗糖的运输和区隔化 蔗糖在细胞质内合成以后的去向主要决定于消耗碳水化合物的器官或组织对蔗糖的需求大小。当需求大时,细胞质内形成的蔗糖优先向这些器官或组织输出,即蔗糖通过某种(些)机理到达质膜或胞间连丝通道进行跨细胞的运输(韧皮部装载)。电镜观察和14C蔗糖示踪研究表明,蔗糖很可能是通过内质网运输的。用呼吸抑制剂和氧化磷酸化解偶联剂抑制细胞质环流时降低蔗糖运输速率,而用ATP处理提高细胞质环流时则可提高蔗糖运输速率,这说明细胞质环流参与了蔗糖的胞内运输,而且这种运输需要呼吸提供能量。 当对蔗糖需求降低时,如去库处理,细胞质内形成的蔗糖则通过跨膜运输进入液泡作为临时性贮藏。蔗糖进入液泡是一种主动过程,因为呼吸抑制剂CCCP(解偶联剂)抑制该过程,而ATP则促进其进行。许多结果表明,蔗糖进入液泡是由位于液泡膜上的蔗糖载体介导的逆蔗糖浓度梯度(图6-17)。 |
| 二、库细胞中同化物的相互转化 蔗糖从库端韧皮部SE-CC复合体卸出,进入库细胞后就可在相应酶的作用下形成淀粉。蔗糖在库细胞内代谢的快慢,不但决定了淀粉形成的速度,收获器官的产量,而且对源端的光合作用及胞内同化物的运输和分配,乃至同化物韧皮部运输(包括装载、长距离运输和卸出)都具有调节作用。 (一)蔗糖的代谢 上述的光合细胞中的蔗糖代谢主要是合成,而库细胞内蔗糖代谢主要是降解。 1.降解代谢 (1)催化蔗糖降解的酶 催化蔗糖降解代谢的酶有两类,一类是转化酶(invertase),另一类是蔗糖合成酶(sucrose synthase SS)。 转化酶 : 它催化蔗糖的水解反应: 蔗糖 H2O→葡萄糖 果糖 蔗糖的旋光性为右旋,而水解后的葡萄糖和果糖混合物的旋光性则转化为左旋,这样的糖称为转化糖,催化此反应的酶就叫转化酶。 根据催化反应所需的最适pH,可将转化酶进一步分成两种,一种称为酸性转化酶,即其催化反应的最适pH为4~5.5,因而该酶只能在酸性环境如质外体和液泡中发挥作用,它主要分布在液泡和细胞壁中。该酶对底物蔗糖的亲和力较高,其Km为2~6mmol·L-1,远低于蔗糖的生理学浓度,而且它的催化活性会受其产物果糖的抑制。另外,酸性转化酶还能被一些可溶性蛋白质所活化。另一类转化酶称为碱性或中性转化酶,其最适pH为7~8,对蔗糖的亲和力相对较低,其Km为10~30mmol·L-1。该酶主要分布在细胞质部分,其生理作用主要有三方面:一是在以蔗糖为最终贮藏物的库器官内起作用;二是在一些缺少酸性转化酶的库组织中催化蔗糖水解,同时提供能量;三是对控制库细胞内己糖水平起重要作用。 ②蔗糖合成酶 它催化下列可逆反应: UDPG 果糖→蔗糖 UDP 该反应的平衡常数为1.3~2。该酶在分解方向的Km(蔗糖)相对较高(30~150mmol·L-1 ),所以细胞中高的蔗糖浓度有利于反应向分解方向进行。
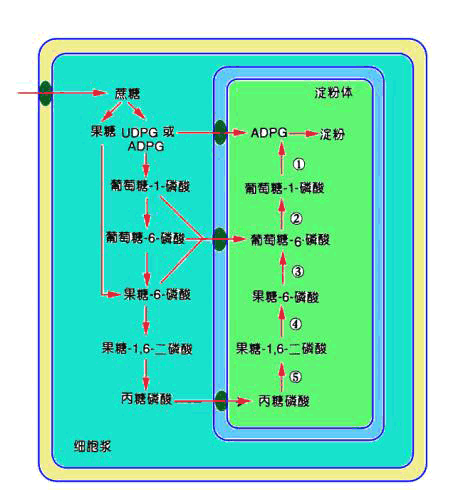
图 6-19 在植物库细胞中的蔗糖代谢和利用途径 A.蔗糖载体; B.己糖或磷酸己糖运转器。 ①转化酶; ②蔗糖合成酶; ③己糖激酶; ④淀粉酶; ⑤磷酸己糖变位酶;⑥UDPG焦磷酸化酶; ⑦ADPG焦磷酸化酶; ⑧淀粉合成酶 蔗糖合成酶位于细胞质内,由4个亚基构成。该酶催化蔗糖分解的活性会被果糖抑制,Mg2 能促进蔗糖的合成,而抑制蔗糖的分解。另外,植株内蔗糖合成酶的活性表现出明显的日变化,即进入光期后,酶的活性开始增加,约在光照6h后达到最大活性。 (2)蔗糖降解途径 蔗糖进入库细胞后由哪一种酶催化降解,取决于植物的种类和库的特性,但两者并不相互排斥。蔗糖降解的途径见图6-19。 当蔗糖由转化酶催化水解时,两反应的产物必须先经过磷酸化才能参与进一步的代谢。而由蔗糖合成酶催化的反应中只有果糖需磷酸化。己糖磷酸化是由己糖激酶催化的,激酶分为两类:一类称己糖激酶,它作用的底物既可以是葡萄糖也可以是果糖,但前者是最适底物。此酶主要存在于由转化酶催化的蔗糖降解的组织或细胞中。另一类为果糖激酶,可催化果糖的磷酸化,该酶存在于由蔗糖合成酶催化的蔗糖降解的组织中。
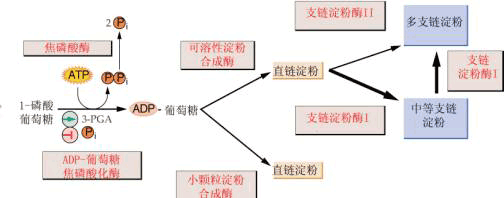
形成的G6P或者进入戊糖磷酸途径,或者由磷酸葡萄糖异构酶催化转变为F6P,或者由磷酸葡萄糖变位酶催化转变为G1P。其中,F6P或者进入糖酵解途径,或者用于合成蔗糖;而G1P是植物体内碳水化合物代谢的关键性中间产物,它可作为由ADPG焦磷酸化酶或UDPG焦磷酸化酶所催化反应的底物。 2.合成代谢 以蔗糖为最终贮藏物的植物和器官,如甘蔗、甜菜块根等能进行蔗糖的合成反应。在这些植物的库器官中具有催化蔗糖合成的酶类,如蔗糖磷酸合成酶、6-磷酸蔗糖磷酸酯酶等。这些酶的催化特性、调节机理与源器官中的类似。另外,韧皮部运输的碳水化合物都可以通过不同的途径成为库器官蔗糖合成的原料(底物)。运输的蔗糖可通过“降解-再合成”(breakdownresynthesis)的方式合成蔗糖,即蔗糖经过韧皮部卸出而进入库细胞,在转化酶或蔗糖合成酶作用下,转化为(磷酸)己糖,然后,再在催化蔗糖合成酶的作用下合成蔗糖。 (二)淀粉的合成 淀粉是光合作用的终产物之一,也是许多农作物收获器官的主要贮藏性多糖之一。由于光合细胞和库组织淀粉合成途径相似,本处仅讨论淀粉合成的一般途径和调节机理。 1.淀粉的合成途径和部位 催化淀粉合成的途径有两条,一条称ADP葡萄糖(ADPG)途径; 另一条为淀粉磷酸化酶催化的途径。然而,植物体内淀粉磷酸化酶主要催化淀粉降解代谢。因此,这里主要介绍ADPG途径。 (1)催化淀粉合成的酶 催化淀粉合成的关键性酶有下列三种:
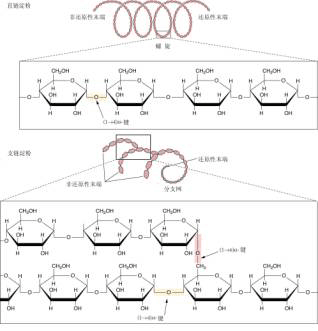
显示细胞质代谢物进入淀粉体的三条可能途径。 A.腺苷酸转运器; B.己糖或磷酸己糖转运器; C.磷酸丙糖转运器; ST.蔗糖载体。 ①ADPG焦磷酸化酶; ②磷酸葡萄糖变位酶; ③己糖异构酶; ④果糖-1,6-二磷酸酯 酶; ⑤醛缩酶 ADPG焦磷酸化酶(ADP glucose pyrophosphorylase AGP) 催化葡萄糖供体ADPG的合成: G1P ATP→ADPG PPi 淀粉合成酶(starch synthase) 催化ADPG分子中的葡萄糖转移到α-1,4-葡聚糖引物的非还原性末端: ADPG (葡聚糖)n→ADP (葡聚糖) 分支酶(branching enzyme) 催化α-1,6-糖苷键的形成: 线性葡聚糖链→具α-1,6-糖苷键的分支葡聚糖链 分支酶对淀粉合成具有两方面意义:其一,使淀粉的α-1,4-糖苷键连接的直链变为含有α-1,6-糖苷键连接的支链,使葡聚糖的分子量不断增大,以便让有限的细胞空间能容纳更多的具有能量的物质;其二,α-1,6糖苷键的导入使葡聚糖的非还原性末端增加,这有利于ADPG焦磷酸化酶和淀粉合成酶的催化反应,使它们能在短时间内催化合成更多的淀粉。
2.调节 有关淀粉合成的调节,目前主要集中在对催化ADPG合成的关键性酶AGP的研究上。所有的证据表明,3PGA是AGP的主要活化剂,而Pi是主要的抑制剂。3PGA还提高了酶对其底物G1P和ATP的亲和力,同时降低对Pi的敏感性。进一步研究表明,调节AGP活性的并不是3PGA或Pi的绝对量,而是3PGA/Pi的比值。 |