
生物膜(biomembrane)是指构成细胞的所有膜的总称。按其所处位置可分为两种:一种处于细胞质外面的一层膜叫质膜,也可叫原生质膜;另一种是处于细胞质中构成各种细胞器的膜,叫内膜(endomembrane)。质膜可由内膜转化而来(如子细胞的质膜由高尔基体小泡融合而成)。
生物膜是细胞结构的基本形式,它对酶催化反应的有序进行和整个细胞的区域化都提供了一个必要的结构基础。当然,生物膜的功能是多种多样的,如细胞的物质代谢、能量转换和信息传递等都与生物膜有关。
| 一、生物膜的化学组成 在真核细胞中,膜结构占整个细胞干重的70%~80%。生物膜由蛋白质、脂类、糖、水和无机离子等组成。蛋白质约占60%~65%,脂类占25%~40%,糖占5%。这些组分,尤其是脂类与蛋白质的比例,因不同细胞、细胞器或膜层而相差很大。功能复杂的膜,其蛋白质含量可达80%,而有的只占20%左右。需说明的是,由于脂类分子的体积比蛋白质分子的小得多,因此生物膜中的脂类分子的数目总是远多于蛋白质分子的数目。如在一个含50%蛋白质的膜中,大概脂类分子与蛋白质分子的比为50∶1。 这一比例关系反映到生物膜结构上,就是脂类以双分子层构成生物膜的基本结构,而蛋白质分子则“镶嵌”于其中。 (一)膜蛋白 生物膜中的蛋白质约占细胞蛋白总量的20%~30%,它们或是单纯的蛋白质,或是与糖、脂结合形成的结合蛋白。根据它们与膜脂相互作用的方式及其在膜中的排列部位,可以大体地将膜蛋白分为两类:外在蛋白与内在蛋白(图1-7)。外在蛋白(extrinsic protein)为水溶性球状蛋白质,通过静电作用及离子键等非共价键与膜脂相连,分布在膜的内外表面。内在蛋白(intrinsic protein)占膜蛋白总量的70%~80%,又叫嵌入蛋白或整合蛋白,其主要特征是水不溶性,分布在脂质双分子层中,有的横跨全膜也称跨膜蛋白(transmembrane protein),有的全部埋入疏水区,有的与外在蛋白结合以多酶复合体形式与膜脂结合。最近,又在生物中发现一类新的膜蛋白,叫膜脂蛋白,它们的蛋白部分不直接嵌入膜,而依赖所含的脂肪酸插入脂质双分子层中。 膜蛋白执行着生物膜的主要功能。不同生物膜所具有的不同生物学功能主要是由于所含膜蛋白的种类和数量的不同。
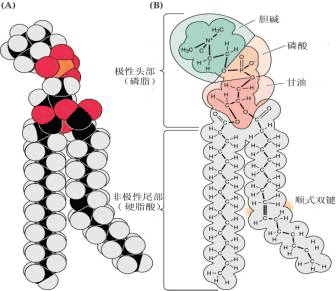
(二)膜脂 在植物细胞中,构成生物膜的脂类主要是复合脂类(complex lipids),包括磷脂、糖脂、硫脂等。 磷脂(phospholipid) 是含磷酸基的复合脂。在植物细胞膜中重要的磷脂属甘油磷脂,它们是磷脂酰胆碱(卵磷脂,phosphatidylcholine)和磷脂酰乙醇胺(脑磷脂,phosphatidylethanolamine)。另外,还有磷脂酰丝氨酸(phosphatidylserine),磷脂酰甘油(phosphatidylglycerinele),磷脂酰肌醇(phosphatidylinositol)等。 磷脂分子结构既有疏水基团,又有亲水基团。
由上可知,膜上的脂类几乎都是两性分子,在水相中可自发地形成脂双层,即脂类分子呈两层排列,亲水的头部处于水相,疏水的尾部朝向中央。这种自发的排列过程称作脂类的自我装配。脂双层一旦有破损也能自我闭合。实验表明:脂双层是流动的,脂类分子能在各自的单分子层内迅速地移动,即横向扩散,而一般不容易“翻转”,即不易从一个单分子层颠转到另一单分子层。脂双层的自我装配、自我闭合以及具有流动性的三大特点决定了它能成为生物膜理想的基本结构。 膜脂上的脂肪酸(fatty acid)有饱和脂肪酸和不饱和脂肪酸之分,不饱和脂肪酸分子有双键,其顺式和反式的互变使不饱和脂肪酸易于弯曲或转动,从而使得膜结构比较松散而不僵硬。 膜脂上的不饱和脂肪酸与植物的抗逆性有很大关系,通常耐寒性强的植物,其膜脂中不饱和脂肪酸含量较高,而且不饱和程度(双键数目)也较高,有利于保持膜在低温时的流动性;而抗热性强的植物,其饱和脂肪酸的含量较高,有利于保持膜在高温时的稳定性。 (三)膜糖 生物膜中的糖类主要分布于质膜的外单分子层。这些糖是不超过15个单糖残基所连接成的具分支的低聚糖链(寡糖链),它们大多数与膜蛋白共价结合,少部分与膜脂结合,分别形成糖蛋白和糖脂(图1-7)。由于单糖彼此间结合方式、排列顺序、种类、数量以及有无分支等差别,其组合是千变万化的,所形成的寡糖链种类非常多,形成了多种细胞表面特异的图像,细胞之间借此进行互相识别和交换信息。 |
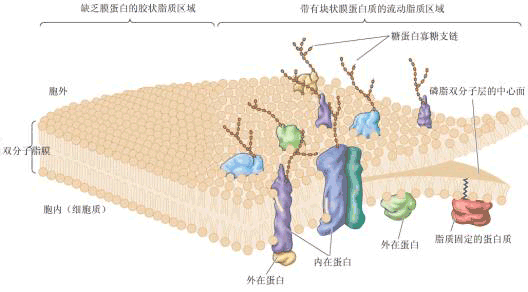
| 二、生物膜的结构 关于生物膜的结构有许多假说与模型,下面介绍两种模型。 (一)流动镶嵌模型 流动镶嵌模型(fluid mosaic model)由辛格尔(S.J. Singer)和尼柯尔森(G. Nicolson)在1972年提出,认为液态的脂质双分子层中镶嵌着可移动的蛋白质,图1-7展示了此模型的结构特点。内在蛋白嵌合在磷脂分子层中,内在蛋白或其聚合体可横穿膜层,两端极性部分伸向水相,中间疏水部分与脂肪酸部分呈疏水结合,外在蛋白与膜两侧的极性部分结合。 这个模型的特点是强调膜的不对称性和流动性。不对称性主要是由脂类和蛋白质分布的不对称造成的。虽然同一种磷脂可见于脂双层的任一层,但它们的数量是不等的。蛋白质在膜中有的半埋于内分子层,有的半埋于外分子层,即使贯穿全膜的蛋白质也是不对称的。另外,寡糖链的分布也是不对称的,它们大多分布于外分子层。膜的流动性包含两个方面,其一是脂类分子是液晶态可动的,脂类分子随温度改变经常处于液晶态和液态的动态平衡之中,两相中脂类分子排列不同,流动性大小也不同。其二是分布于膜脂双分子层的蛋白质也是流动的,它们可以在脂分子层中侧向扩散,但不能翻转扩散。这说明了少量膜脂与膜蛋白有相对专一的作用,这种作用是膜蛋白行使功能所必须的。 脂双层的流动性保证了膜能经受一定程度的形变而不致破裂,这可使膜中各种成分按需要调整或组合,使之合理分布,有利于表现膜的多种功能。更重要的是它允许膜互相融合而不失去膜对通透性的控制,确保膜分子在细胞分裂时均等地分配给子代细胞。如果膜不具有流动性,则很难想象细胞如何存活、生长和繁殖。流动镶嵌模型虽得到比较广泛的支持,但仍有很多局限性,如忽视了蛋白质对脂类分子流动性的控制作用和膜各部分流动的不均匀性等问题。 (二)板块镶嵌模型 板块镶嵌模型(plate mosaic model)由贾因(M.K. Gain)和怀特(White)在1977年提出。他们认为,由于生物膜脂质可以在环境温度或其它化学成分变化的影响下,或是由于膜中同时存在着不同脂质(脂肪链的长短或不同的饱和度),或者由于蛋白质和蛋白质、蛋白质和脂质间的相互作用,使膜脂的局部经常处于一种“相变”状态,即一部分脂区表现为从液晶态转变为晶态,而另一部分脂区表现为从晶态转变为液晶态。因此,整个生物膜可以看成是由不同组织结构、不同大小、不同性质、不同流动性的可移动的“板块”所组成,高度流动性的区域和流动性比较小的区域可以同时存在,随着生理状态和环境条件的改变,这些“板块”之间可以彼此转化。 |
| 三、生物膜的功能 在生命起源的最初阶段,正是有了脂性的膜,才使生命物质——蛋白质与核酸获得与周围介质隔离的屏障而保持聚集和相对稳定的状态,继之才有细胞的发展。因此,质膜是任何活细胞必不可少的。植物细胞可以脱离细胞壁而生活,却不能脱离质膜而生存。膜的主要功能如下: 1.分室作用 细胞的膜系统不仅把细胞与外界环境隔开,而且把细胞内的空间分隔,使细胞内部的区域化(compartmentation),即形成各种细胞器,从而使细胞的代谢活动“按室进行”。各区域内均具特定的pH、电位、离子强度和酶系等。 同时,由于内膜系统的存在,又将各个细胞器联系起来共同完成各种连续的生理生化反应,比如光呼吸过程就是由叶绿体、过氧化物体和线粒体三者协同完成的。 2.代谢反应的场所 细胞内的许多生理生化过程在膜上有序进行。如光合作用的光能吸收、电子传递和光合磷酸化、呼吸作用的电子传递及氧化磷酸化过程分别是在叶绿体的光合膜和线粒体内膜上进行的。 3.物质交换 质膜的另一个重要特性是对物质的透过具有选择性,控制膜内外进行物质交换。如质膜可通过扩散、离子通道、主动运输及内吞外排等方式来控制物质进出细胞。各种细胞器上的膜也通过类似方式控制其小区域与胞质进行物质交换。 4.识别功能 质膜上的多糖链分布于其外表面,似“触角”一样能够识别外界物质,并可接受外界的某种刺激或信号,使细胞作出相应的反应。例如,花粉粒外壁的糖蛋白与柱头细胞质膜的蛋白质之间就可进行识别反应。膜上还存在着各种各样的受体(receptor),能感应刺激,传导信息,调控代谢。 |
生物膜(biomembrane)是指构成细胞的所有膜的总称。按其所处位置可分为两种:一种处于细胞质外面的一层膜叫质膜,也可叫原生质膜;另一种是处于细胞质中构成各种细胞器的膜,叫内膜(endomembrane)。质膜可由内膜转化而来(如子细胞的质膜由高尔基体小泡融合而成)。
生物膜是细胞结构的基本形式,它对酶催化反应的有序进行和整个细胞的区域化都提供了一个必要的结构基础。当然,生物膜的功能是多种多样的,如细胞的物质代谢、能量转换和信息传递等都与生物膜有关。
| 一、生物膜的化学组成 在真核细胞中,膜结构占整个细胞干重的70%~80%。生物膜由蛋白质、脂类、糖、水和无机离子等组成。蛋白质约占60%~65%,脂类占25%~40%,糖占5%。这些组分,尤其是脂类与蛋白质的比例,因不同细胞、细胞器或膜层而相差很大。功能复杂的膜,其蛋白质含量可达80%,而有的只占20%左右。需说明的是,由于脂类分子的体积比蛋白质分子的小得多,因此生物膜中的脂类分子的数目总是远多于蛋白质分子的数目。如在一个含50%蛋白质的膜中,大概脂类分子与蛋白质分子的比为50∶1。 这一比例关系反映到生物膜结构上,就是脂类以双分子层构成生物膜的基本结构,而蛋白质分子则“镶嵌”于其中。 (一)膜蛋白 生物膜中的蛋白质约占细胞蛋白总量的20%~30%,它们或是单纯的蛋白质,或是与糖、脂结合形成的结合蛋白。根据它们与膜脂相互作用的方式及其在膜中的排列部位,可以大体地将膜蛋白分为两类:外在蛋白与内在蛋白(图1-7)。外在蛋白(extrinsic protein)为水溶性球状蛋白质,通过静电作用及离子键等非共价键与膜脂相连,分布在膜的内外表面。内在蛋白(intrinsic protein)占膜蛋白总量的70%~80%,又叫嵌入蛋白或整合蛋白,其主要特征是水不溶性,分布在脂质双分子层中,有的横跨全膜也称跨膜蛋白(transmembrane protein),有的全部埋入疏水区,有的与外在蛋白结合以多酶复合体形式与膜脂结合。最近,又在生物中发现一类新的膜蛋白,叫膜脂蛋白,它们的蛋白部分不直接嵌入膜,而依赖所含的脂肪酸插入脂质双分子层中。 膜蛋白执行着生物膜的主要功能。不同生物膜所具有的不同生物学功能主要是由于所含膜蛋白的种类和数量的不同。
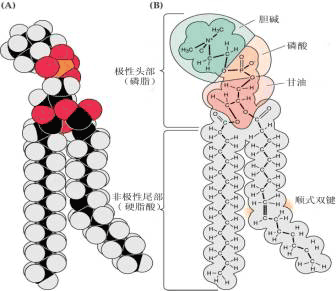
(二)膜脂 在植物细胞中,构成生物膜的脂类主要是复合脂类(complex lipids),包括磷脂、糖脂、硫脂等。 磷脂(phospholipid) 是含磷酸基的复合脂。在植物细胞膜中重要的磷脂属甘油磷脂,它们是磷脂酰胆碱(卵磷脂,phosphatidylcholine)和磷脂酰乙醇胺(脑磷脂,phosphatidylethanolamine)。另外,还有磷脂酰丝氨酸(phosphatidylserine),磷脂酰甘油(phosphatidylglycerinele),磷脂酰肌醇(phosphatidylinositol)等。 磷脂分子结构既有疏水基团,又有亲水基团。
由上可知,膜上的脂类几乎都是两性分子,在水相中可自发地形成脂双层,即脂类分子呈两层排列,亲水的头部处于水相,疏水的尾部朝向中央。这种自发的排列过程称作脂类的自我装配。脂双层一旦有破损也能自我闭合。实验表明:脂双层是流动的,脂类分子能在各自的单分子层内迅速地移动,即横向扩散,而一般不容易“翻转”,即不易从一个单分子层颠转到另一单分子层。脂双层的自我装配、自我闭合以及具有流动性的三大特点决定了它能成为生物膜理想的基本结构。 膜脂上的脂肪酸(fatty acid)有饱和脂肪酸和不饱和脂肪酸之分,不饱和脂肪酸分子有双键,其顺式和反式的互变使不饱和脂肪酸易于弯曲或转动,从而使得膜结构比较松散而不僵硬。 膜脂上的不饱和脂肪酸与植物的抗逆性有很大关系,通常耐寒性强的植物,其膜脂中不饱和脂肪酸含量较高,而且不饱和程度(双键数目)也较高,有利于保持膜在低温时的流动性;而抗热性强的植物,其饱和脂肪酸的含量较高,有利于保持膜在高温时的稳定性。 (三)膜糖 生物膜中的糖类主要分布于质膜的外单分子层。这些糖是不超过15个单糖残基所连接成的具分支的低聚糖链(寡糖链),它们大多数与膜蛋白共价结合,少部分与膜脂结合,分别形成糖蛋白和糖脂(图1-7)。由于单糖彼此间结合方式、排列顺序、种类、数量以及有无分支等差别,其组合是千变万化的,所形成的寡糖链种类非常多,形成了多种细胞表面特异的图像,细胞之间借此进行互相识别和交换信息。 |
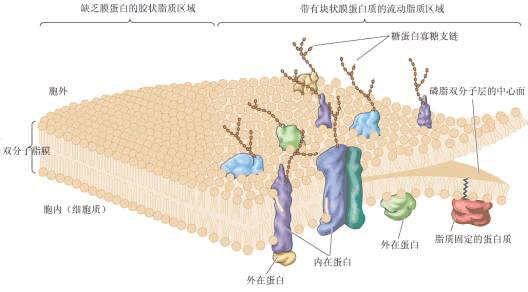
| 二、生物膜的结构 关于生物膜的结构有许多假说与模型,下面介绍两种模型。 (一)流动镶嵌模型 流动镶嵌模型(fluid mosaic model)由辛格尔(S.J. Singer)和尼柯尔森(G. Nicolson)在1972年提出,认为液态的脂质双分子层中镶嵌着可移动的蛋白质,图1-7展示了此模型的结构特点。内在蛋白嵌合在磷脂分子层中,内在蛋白或其聚合体可横穿膜层,两端极性部分伸向水相,中间疏水部分与脂肪酸部分呈疏水结合,外在蛋白与膜两侧的极性部分结合。 这个模型的特点是强调膜的不对称性和流动性。不对称性主要是由脂类和蛋白质分布的不对称造成的。虽然同一种磷脂可见于脂双层的任一层,但它们的数量是不等的。蛋白质在膜中有的半埋于内分子层,有的半埋于外分子层,即使贯穿全膜的蛋白质也是不对称的。另外,寡糖链的分布也是不对称的,它们大多分布于外分子层。膜的流动性包含两个方面,其一是脂类分子是液晶态可动的,脂类分子随温度改变经常处于液晶态和液态的动态平衡之中,两相中脂类分子排列不同,流动性大小也不同。其二是分布于膜脂双分子层的蛋白质也是流动的,它们可以在脂分子层中侧向扩散,但不能翻转扩散。这说明了少量膜脂与膜蛋白有相对专一的作用,这种作用是膜蛋白行使功能所必须的。 脂双层的流动性保证了膜能经受一定程度的形变而不致破裂,这可使膜中各种成分按需要调整或组合,使之合理分布,有利于表现膜的多种功能。更重要的是它允许膜互相融合而不失去膜对通透性的控制,确保膜分子在细胞分裂时均等地分配给子代细胞。如果膜不具有流动性,则很难想象细胞如何存活、生长和繁殖。流动镶嵌模型虽得到比较广泛的支持,但仍有很多局限性,如忽视了蛋白质对脂类分子流动性的控制作用和膜各部分流动的不均匀性等问题。 (二)板块镶嵌模型 板块镶嵌模型(plate mosaic model)由贾因(M.K. Gain)和怀特(White)在1977年提出。他们认为,由于生物膜脂质可以在环境温度或其它化学成分变化的影响下,或是由于膜中同时存在着不同脂质(脂肪链的长短或不同的饱和度),或者由于蛋白质和蛋白质、蛋白质和脂质间的相互作用,使膜脂的局部经常处于一种“相变”状态,即一部分脂区表现为从液晶态转变为晶态,而另一部分脂区表现为从晶态转变为液晶态。因此,整个生物膜可以看成是由不同组织结构、不同大小、不同性质、不同流动性的可移动的“板块”所组成,高度流动性的区域和流动性比较小的区域可以同时存在,随着生理状态和环境条件的改变,这些“板块”之间可以彼此转化。 |
三、生物膜的功能
|
