
第四节 植物的蒸腾作用
植物经常处于吸水和失水的动态平衡之中。植物一方面从土壤中吸收水分,另一方面又向大气中蒸发水分。陆生植物在一生中耗水量很大。据估算,一株玉米一生需耗水200kg以上。其中只有极少数(约占1.5%~2%)水分是用于体内物质代谢,绝大多数都散失到体外。其散失的方式,除了少量的水分以液体状态通过吐水的方式散失外,大部分水分则以气态,即以蒸腾作用的方式散失。所谓蒸腾作用(transpiration) 是指植物体内的水分以气态散失到大气中去的过程。与一般的蒸发不同,蒸腾作用是一个生理过程,受到植物体结构和气孔行为的调节。
| 一、 蒸腾作用的生理意义和方式 (一)蒸腾作用的生理意义 陆生植物在进行光合和呼吸的过程中,以伸展在空中的枝叶与周围环境发生气体交换,然而随之而来的是大量地丢失水分。蒸腾作用消耗水分,这对陆生植物来说是不可避免的,它既会引起水分亏缺,破坏植物的水分平衡,甚至引起祸害,但同时,它又对植物的生命活动具有一定的意义。 1.蒸腾作用能产生的蒸腾拉力 蒸腾拉力是植物被动吸水与转运水分的主要动力,这对高大的乔木尤为重要。 2.蒸腾作用促进木质部汁液中物质的运输 土壤中的矿质盐类和根系合成的物质可随着水分的吸收和集流而被运输和分布到植物体各部分去。 3.蒸腾作用能降低植物体的温度 这是因为水的气化热高,在蒸腾过程中可以散失掉大量的辐射热。 4.蒸腾作用的正常进行有利于CO2的同化 这是因为叶片进行蒸腾作用时,气孔是开放的,开放的气孔便成为CO2进入叶片的通道。 (二)蒸腾作用的方式 蒸腾作用有多种方式。幼小的植物,暴露在地上部分的全部表面都能蒸腾。植物长大后,茎枝表面形成木栓,未木栓化的部位有皮孔,可以进行皮孔蒸腾(lenticular transpiration)。但皮孔蒸腾的量甚微,仅占全部蒸腾量的0.1%左右,植物的茎、花、果实等部位的蒸腾量也很有限,因此,植物蒸腾作用绝大部分是靠叶片进行的。 叶片的蒸腾作用方式有两种,一是通过角质层的蒸腾,称为角质蒸腾(cuticular transpiration);二是通过气孔的蒸腾,称为气孔蒸腾(stomatal transpiration)。角质层本身不易让水通过,但角质层中间含有吸水能力强的果胶质,同时角质层也有孔隙,可让水分自由通过。角质层蒸腾和气孔蒸腾在叶片蒸腾中所占的比重,与植物的生态条件和叶片年龄有关,实质上也就是和角质层厚薄有关。例如:阴生和湿生植物的角质蒸腾往往超过气孔蒸腾。幼嫩叶子的角质蒸腾可达总蒸腾量的1/3到1/2。一般植物成熟叶片的角质蒸腾,仅占总蒸腾量的3%~5%。因此,气孔蒸腾是中生和旱生植物蒸腾作用的主要方式。 |
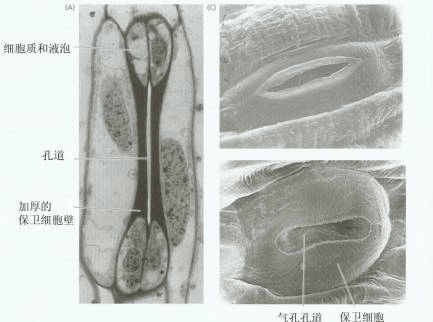
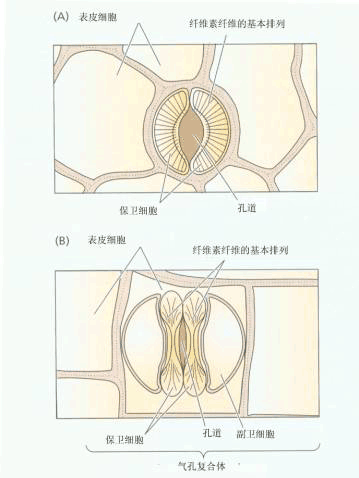
| 二、气孔蒸腾 气孔是植物进行体内外气体交换的重要门户。水蒸气、CO2、O2都要共用气孔这个通道,气孔的开闭会影响植物的蒸腾、光合、呼吸等生理过程。 气孔是植物叶片表皮组织的小孔,一般由成对的保卫细胞(guard cell)组成。保卫细胞四周环绕着表皮细胞,毗连的表皮细胞如在形态上和其它表皮细胞相同,就称之为邻近细胞(neighbouring cell),如有明显区别,则称为副卫细胞(subsidiary cell)。保卫细胞与邻近细胞或副卫细胞构成气孔复合体。保卫细胞在形态上和生理上与表皮细胞有显著的差别。 (一)气孔的形态结构及生理特点 1.气孔数目多、分布广 气孔的大小、数目和分布因植物种类和生长环境而异(表2-3)。一般单子叶植物叶的上下表皮都有气孔分布,而双子叶植物主要分布在下表皮。浮水植物气孔都分布在上表皮。
4.保卫细胞具有多种细胞器 保卫细胞中细胞器的种类比其他表皮细胞中的多,特别是含有较多的叶绿体。保卫细胞中的叶绿体具有光化学活性,能进行光合磷酸化合成ATP,只是缺少固定CO2的关键酶Rubisco,但是保卫细胞的细胞质中含有PEP羧化酶,能进行PEP的羧化反应,其产物为苹果酸(PEP HCO3-→苹果酸)。叶绿体内含有淀粉体,在白天光照下淀粉会减少,而暗中淀粉则积累。这和正常的光合组织中恰好相反。此外,保卫细胞中还含有异常丰富的线粒体,为叶肉细胞的5~10倍,推测其呼吸旺盛,能为开孔时的离子转运提供能量。
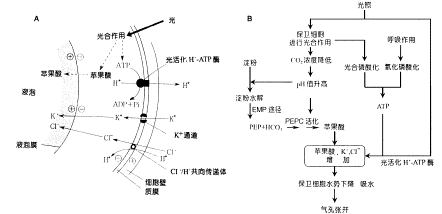
6.保卫细胞与周围细胞联系紧密 保卫细胞与副卫细胞或邻近细胞间没有胞间连丝,相邻细胞的壁很薄,质膜上存在有ATPase、K 通道,另外在保卫细胞外壁上还有外连丝(ectodesmata)结构,它也可作为物质运输的通道。这些结构有利于保卫细胞同副卫细胞或邻近细胞在短时间内进行H 、K 交换,以快速改变细胞水势。而有胞间连丝的细胞,细胞间的水和溶质分子可经胞间连丝相互扩散,不利于二者间建立渗透势梯度。 另外,保卫细胞能感受内、外信号而调节自身体积,从而控制气孔大小,主宰植物体与外界环境间的水分、气体等交换。因此,保卫细胞可说得上是植物体中奇妙的细胞。 (二)气孔运动的机理 气孔运动是由保卫细胞水势的变化而引起的。20世纪70年代以前,人们认为保卫细胞间水势的变化是由细胞中淀粉与葡萄糖的相互转变引起的,曾流行过“淀粉—糖转化学说”,即淀粉转化为葡萄糖时开孔,葡萄糖转化为淀粉时闭孔。以后的实验证明,保卫细胞的水势变化是由K 及苹果酸等渗透调节物质进出保卫细胞引起的(表2-5)。下面介绍有关气孔运动的两种学说。
实验还发现,在K 进入保卫细胞的同时,还伴随着等量负电荷的阴离子进入,以保持保卫细胞的电中性,这也具有降低水势的效果。在暗中,光合作用停止,H ATP酶因得不到所需的ATP而停止做功,从而使保卫细胞的质膜去极化(depolarization),以驱使K 经外向K 通道(outward K channel)向周围细胞转移,并伴随着阴离子的释放,这样导致了保卫细胞水势升高,水分外移,使气孔关闭。在干旱胁迫(drought stress)下,脱落酸含量增加,可通过增加胞质钙浓度,使得保卫细胞的质膜去极化,驱动外向K 通道,促进K 、Cl-流出,同时抑制K 流入,以降低保卫细胞膨压,导致气孔关闭。 2.苹果酸代谢学说(malate metabolism theory) 20世纪70年代初以来,人们发现苹果酸在气孔开闭运动中起着某种作用。在光照下, 保卫细胞内的部分CO2被利用时,pH就上升至8.0~8.5,从而活化了PEP羧化酶,它可催化由淀粉降解产生的PEP与HCO3-结合形成草酰乙酸,并进一步被NADPH还原为苹果酸。
综合上述学说,气孔运动的可能机理可用图2-12来说明。
光照下保卫细胞液泡中的离子积累。由光合作用生成的ATP驱动H 泵,向质膜外泵出H ,建立膜内外的H 梯度,在H 电化学势的驱动下,K 经K 通道、Cl-经共向传递体进入保卫细胞。另外,光合作用生成苹果酸。K 、Cl-和苹果酸进入液泡,降低保卫细胞的水势。B.气孔开启机理图解。 (三)影响气孔运动的因素 气孔运动(stomatal movement)受着内部因素和外界因素的调节。 气孔的特殊结构和生理特性使气孔运动具有调节的功能。以下讨论外界因素对气孔运动的影响。 1.光 光是气孔运动的主要调节因素。光可促进保卫细胞内苹果酸的形成和K 、Cl-的积累。一般情况下,光可促进气孔张开,暗中则气孔关闭。但景天科酸代谢植物例外,它们的气孔通常是白天关闭,夜晚张开。不同植物气孔张开所需光强有所不同,例如烟草只要有完全日照的2.5%光强即可,而大多数植物则要求较高的光强。光促进气孔开启的效应有两种,一种是通过光合作用发生的间接效应,这种效应被光合电子传递抑制剂DCMU所抑制:另一种是通过光受体感受光信号而发生的直接效应,这种效应不被DCMU抑制。红光和蓝光都可引起气孔张开,但蓝光使气孔张开的效率是红光的10倍。通常认为,红光是通过间接效应,而蓝光是直接对气孔开启起作用的。红光的光受体可能是叶绿素;蓝光的光受体可能是隐花色素(cryptochrome)。有人认为蓝光能活化质膜H -ATP酶,不断泵出H ,形成跨膜电化学势梯度,它是K 通过K 通道移动的动力,可使保卫细胞内K 浓度增加,水势降低,气孔张开。 2.二氧化碳 CO2对气孔运动影响很大。低浓度CO2促进气孔张开,高浓度CO2能使气孔迅速关闭(无论光下或暗中都是如此)。在高浓度CO2下,气孔关闭的可能原因是:①高浓度CO2会使质膜透性增加,导致K 泄漏,消除质膜内外的溶质势梯度,②CO2使细胞内酸化,影响跨膜质子浓度差的建立。 3.温度 气孔开度(stomatal aperture)一般随温度的上升而增大。在30℃左右气孔开度最大,超过30℃或低于10℃的低温下,气孔部分张开或关闭。这表明气孔运动是与酶促反应有关的生理过程。 4.水分 气孔运动与保卫细胞膨压变化密切相关, 而膨压变化又是由于水分进出保卫细胞引起的,因此,叶片的水分状况是直接影响气孔运动的关键因素。植物处于水分胁迫条件下气孔开度减小,以减少水分的丢失。如果久雨,表皮细胞为水饱和,挤压保卫细胞,气孔会关闭。如果蒸腾过于强烈,保卫细胞失水过多,即使在光下,气孔也会关闭。
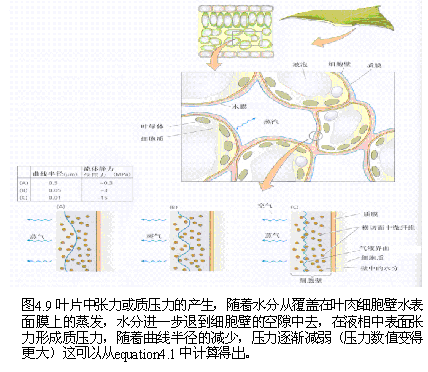
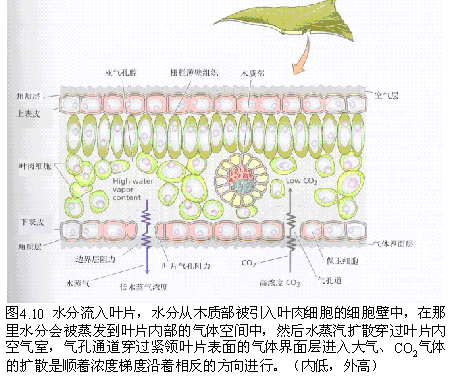
(四)影响气孔蒸腾的内外因素 气孔蒸腾本质上是一个蒸发过程。气孔蒸腾的第一步是位于气孔下腔(substomatal cavities)周围的叶肉细胞的细胞壁中的水分变成水蒸气,然后经过气孔下腔和气孔扩散到叶面的扩散层,再由扩散层扩散到空气中去。蒸腾速率与水蒸气由气孔向外的扩散力成正比,而与扩散途径的阻力成反比。扩散力大小决定于气孔下腔蒸气压与叶外蒸气压之差,即蒸气压梯度(vapor pressure gradient)。 蒸气压差愈大,蒸腾速率愈快;反之,则慢。扩散阻力包括气孔阻力和扩散层阻力,其中气孔阻力主要受气孔开度制约,扩散层阻力主要决定于扩散层的厚薄。气孔阻力大,扩散层厚,蒸腾慢;反之则快。
(1)气孔频度 气孔频度(stomatal frequency)为1mm-2叶片气孔数上的,气孔频度大有利于蒸腾的进行。 (2)气孔大小 气孔孔径较大,内部阻力小,蒸腾较强。 (3)气孔下腔 气孔下腔容积大,叶内外蒸气压差大,蒸腾快。 (4)气孔开度 气孔开度大,蒸腾快,反之,蒸腾减弱。 (5)气孔构造 气孔构造不同也会影响蒸腾作用,气孔下陷的,扩散层相对加厚,阻力大,蒸腾较慢。 2.外界条件对气孔蒸腾的影响 蒸腾速率取决于叶片内外的蒸气压差和扩散途径阻力的大小。所以,凡是影响叶片内外蒸气压差和扩散途径阻力的外界条件,都会影响蒸腾速率的高低。 (1)光照 光对蒸腾作用的影响首先是引起气孔的开放,减少气孔阻力,从而增强蒸腾作用。其次,光可以提高大气与叶子温度,增加叶内外蒸气压差,加快蒸腾速率。 (2)温度 温度对蒸腾速率影响很大。当大气温度升高时,叶温比气温高出2~10℃,因而,气孔下腔蒸气压的增加大于空气蒸气压的增加,这样叶内外蒸气压差加大,蒸腾加强。当气温过高时,叶片过度失水,气孔会关闭,使蒸腾减弱。 (3)湿度 在温度相同时,大气的相对湿度越大,其蒸气压就越大,叶内外蒸气压差就变小,气孔下腔的水蒸气不易扩散出去,蒸腾减弱;反之,大气相对湿度较低,则蒸腾速度加快。 (4)风速 风速较大时,可将叶面气孔外水蒸气扩散层吹散,而代之以相对湿度较低的空气,既减少了扩散阻力,又增大了叶内外蒸气压差,可以加速蒸腾。强风可能会引起气孔关闭或开度减小,内部阻力加大,蒸腾减弱。 蒸腾作用的昼夜变化主要是由外界条件所决定的。以水稻为例,在一天当中,早上7 时开始逐渐增大,到上午10时迅速上升,中午13时左右达到高峰,而下午14时后逐渐下降,18时后则迅速下降。蒸腾强度的这种日变化是与光强和气温变化一致的,特别是与光强的关系更为密切。 |

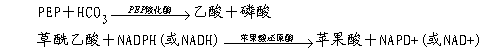
| 三、腾作用的指标和测定方法 (一)蒸腾作用的指标 蒸腾作用的强弱,可以反映出植物体内水分代谢的状况或植物对水分利用的效率。蒸腾作用常用的指标有: 1.蒸腾速率(transpiration rate) 又称蒸腾强度或蒸腾率。指植物在单位时间内、单位叶面积上通过蒸腾作用散失的水量。常用单位:g·m-2·h-1、mg·dm-2·h-1。大多数植物白天的蒸腾速率是15~250g·m-2·h-1,夜晚是1~20g·m-2·h-1。 2.蒸腾效率(transpiration ratio) 指植物每蒸腾1kg水时所形成的干物质的g数。常用单位:g·kg-1。一般植物的蒸腾效率为1~8g·kg-1。 3.蒸腾系数(transpiration coefficient) 又称需水量(water requirement)。指植物每制造1g干物质所消耗水分的g数,它是蒸腾效率的倒数。大多数植物的蒸腾系数在125~1000之间。木本植物的蒸腾系数比较低,白蜡树约85,松树约40;草本植物蒸腾系数较高,玉米为370,小麦为540。蒸腾系数越小,则表示该植物利用水分的效率越高。 (二)蒸腾速率的测定方法 1.植物离体部分的快速称重法 切取植物体的一部分(叶、苗、枝或整个地上部分)迅速称重,2~3min后再次称重,两次重量差即为单位时间内的蒸腾失水量。这个方法的依据是植物离体部分在切割后开始的2~5min内,原有的蒸腾速率无多大改变。称重时可采用扭力天平或电子分析天平。 2.测量重量法 把植株栽在容器中,茎叶外露进行蒸腾作用,容器口适当密封,使容器内的水分不发生散失。在一定间隔的时间里,用电子天平称得容器及植株重量的变化,就可以得到蒸腾速率。 3.量计测定法 这是一种适合于田间条件下测定瞬时蒸腾速率的方法。主要是应用灵敏的湿度敏感元件测定蒸腾室内的空气相对湿度的短期变化。当植物的枝、叶或整株植物放入蒸腾室后,在第一个30s内每隔10s测定一次室内的湿度。蒸腾速率是由绝对湿度增加而得到的,而绝对湿度是由相对湿度的变化速率和同一时刻的空气温度计算出来的。 近年来,有一种稳态气孔计,其透明小室的直径仅1~2cm,将叶片夹在小室间,在微电脑控制下向小室内通入干燥空气,流速恰好能使小室内的湿度保持恒定。然后可根据干燥空气流量的大小计算出蒸腾速率。 4.红外线分析仪测定法 红外线对双元素组成的气体有强烈的吸收能力。H2O是双元素(H和O)组成子,因此,用红外线分析仪(IRGA)可测定水的浓度,即湿度,并用来计算蒸腾速率。 这种仪器是测定两种空气流中水浓度(绝对湿度)的差值,且可以作两种类型的测量:一是绝对测量,即测定蒸腾室中水蒸气浓度与封闭在参比管内的隋性气体或含有已知浓度的水蒸气的浓度的差值。二是相对测量,即测定流入蒸腾室前和流出蒸腾室后的两种水蒸气浓度间的差值。 |
| 四、当降低蒸腾的途径 植物通过蒸腾作用会散失大量的水分,一旦水分供应不足,植物就发生萎蔫。因此,在农业生产上,为了维持作物体内的水分平衡(water balance),就要“开源节流”。除了采取有效措施,促使根系发达,以保证水分供应之外,适当减少蒸腾消耗也是必要的。其途径主要有: 1. 减少蒸腾面积 在移栽植物时,可去掉一些枝叶,减少蒸腾面积, 降低蒸腾失水量,以维持移栽植物体内水分平衡,有利其成活。 2. 降低蒸腾速率 避开促进蒸腾的外界条件,在午后或阴天移栽植物,或栽后搭棚遮荫,这样就能降低移栽植株的蒸腾速率。此外,实行设施栽培,也能降低棚内作物的蒸腾速率,这是由于在密闭的大棚或温室内,相对湿度较高的缘故。 3.使用抗蒸腾剂某些能降低植物蒸腾速率而对光合作用和生长影响不太大的物质,称为抗蒸腾剂(antitranspirant)。按其性质和作用方式不同,可将抗蒸腾剂分为三类: (1)代谢型抗蒸腾剂 这类药物中有些能影响保卫细胞的膨胀,减小气孔开度,如阿特拉津等;也有些能改变保卫细胞膜透性,使水分不易向外扩散,如苯汞乙酸、烯基琥珀酸等。 (2)薄膜型抗蒸腾剂 这类药物施用于植物叶面后能形成单分子薄层,阻碍水分散失,如硅酮、丁二烯丙烯酸等。 (3)反射型抗蒸腾剂 这类药物能反射光,其施用于叶面后,叶面对光的反射增加,从而降低叶温,减少蒸腾量,如高岭土。 |
