植物在整个生长过程中,受到各种内外因素的影响,这就需要植物体正确地辨别各种信息并作出相应的反应,以确保正常的生长和发育。例如植物的向光性能促使植物向光线充足的方向生长,在这个过程中,首先植物体要能感受到光线,然后把相关的信息传递到有关的靶细胞,并诱发胞内信号转导,调节基因的表达或改变酶的活性,从而使细胞作出反应。这种信息的胞间传递和胞内转导过程称为植物体内的信号传导。
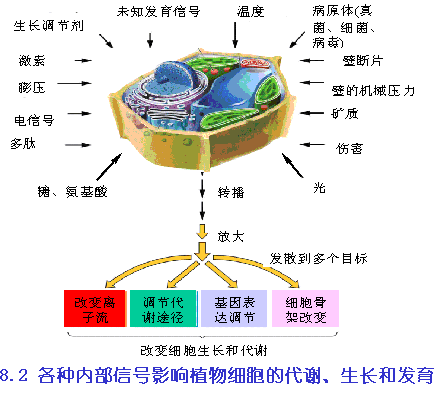 |
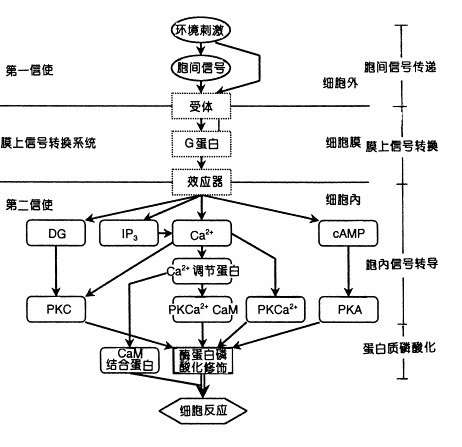
在植物细胞的信号反应中,已发现有几十种信号分子。按其作用范围可分为胞间信号分子和胞内信号分子。对于细胞信号传导的分子途径,可分为四个阶段,即:胞间信号传递、膜上信号转换、胞内信号转导及蛋白质可逆磷酸化(图6-25)。
| 一、胞间信号的传递 植物体内的胞间信号可分为两类,即化学信号和物理信号。 (一)化学信号 化学信号(chemical signal)是指细胞感受刺激后合成并传递到作用部位引起生理反应的化学物质。一般认为,植物激素是植物体主要的胞间化学信号。如当植物根系受到水分亏缺胁迫时,根系细胞迅速合成脱落酸(ABA),ABA再通过木质部蒸腾流输送到地上部分,引起叶片生长受抑和气孔导度的下降。而且ABA的合成和输出量也随水分胁迫程度的加剧而显著增加。这种随着刺激强度的增加,细胞合成量及向作用位点输出量也随之增加的化学信号物质称之为正化学信号(positive chemical signal)。然而在水分胁迫时,根系合成和输出细胞分裂素(CTK)的量显著减少,这样的随着刺激强度的增加,细胞合成量及向作用位点输出量随之减少的化学信号物质称为负化学信号(negative chemical signal)。
图 6-25 细胞信号传导的主要分子途径 IP3.三磷酸肌醇; DG.二酰甘油; PKA.依赖cAMP的蛋白激酶;PK Ca2 依赖Ca2 的蛋白激酶;PKC.依赖Ca2 与磷脂的蛋白激酶;PK Ca2 ·CaM. 依赖Ca2 ·CaM的蛋白激酶 当植物的一张叶片被虫咬伤后,会诱导本叶和其它叶产生蛋白酶抑制物(PIs)等,以阻碍病原菌或害虫进一步侵害。如伤害后立即除去受害叶,则其它叶片不会产生PIs。但如果将受害叶的细胞壁水解片段(主要是寡聚糖)加到叶片中,又可模拟伤害反应诱导PIs的产生,从而认为寡聚糖是由受伤叶片释放并经维管束转移,继而诱导PIs基因活化的信号物质。已知1,3-β-D-葡聚糖、寡聚半乳糖醛酸、富含甘露糖的糖蛋白、聚氨基葡萄糖等都是构成细胞壁的主要成分,它们除了具有支持细胞框架的功能外,还起诱导抗性和控制发育的信号作用,成为引人注目的胞间信号分子。此外,一些生长调节物质如壳梭孢菌素、水杨酸、花生四烯酸、茉莉酸、茉莉酸甲酯、多胺类物质以及乙酰胆碱等都具有化学信号的功能。 (二)物理信号 物理信号(physical signal)是指细胞感受到刺激后产生的能够起传递信息作用的电信号和水力学信号。电信号传递是植物体内长距离传递信息的一种重要方式,是植物体对外部刺激的最初反应。我国著名的植物生理学家娄成后教授在20世纪60年代就指出:“电波的信息传递在高等植物中是普遍存在的。”他认为植物为了对环境变化作出反应,既需要专一的化学信号传递,也需要快速的电波传递。植物的电波研究较多的为动作电位 (action potential,AP),也叫动作电波,它是指细胞和组织中发生的相对于空间和时间的快速变化的一类生物电位。植物中动作电波的传递仅用短暂的冲击(如机械震击、电脉冲或局部温度的升降)就可以激发出来,而且受刺激的植物没有伤害,不久便恢复原状。有人测到捕虫植物的动作电位幅度为110~115mV,传递速度可达6~30cm·s-1。 一些敏感植物或组织(如含羞草的茎叶、攀缘植物的卷须等),当受到外界刺激,发生运动反应(如小叶闭合下垂、卷须弯曲等)时伴有电波的传递。当让平行排列的轮藻细胞中的一个细胞接受电刺激而引起动作电位后,该细胞就可以将其传递到相距10mm处的另一个细胞而且引起同步节奏的动作电位。怀尔登(Wildon)等用番茄做实验,指出由子叶伤害而引起第一真叶产生PIs的过程中,动作电位是传播的主要方式。他们采取让电信号通过后马上就除去子叶以及使子叶叶柄致冷以阻碍筛管运输、排除化学物质传递的试验,其结果都证明单有电信号就可以引起PIs反应,而且他们也首次证明了电信号可引起包括基因转录在内的生理生化变化。 植物细胞对水力学信号(压力势的变化)很敏感。玉米叶片木质部压力的微小变化就能迅速影响叶片气孔的开度,即压力势降低时气孔开放,反之亦然。 (三)胞间信号的传递 当环境信号刺激的作用位点与效应位点处在植物不同部位时,胞间信号就要作长距离的传递,高等植物胞间信号的长距离传递,主要有以下几种。 1.易挥发性化学信号在体内气相的传递 易挥发性化学信号可通过在植株体内的气腔网络 (air space network) 中的扩散而迅速传递,通常这种信号的传递速度可达2mm·s-1左右。植物激素乙烯和茉莉酸甲酯(JA-Me)均属此类信号,而且这两类化合物在植物某器官或组织受到刺激后可迅速合成。在大多数情况下,这些化合物从合成位点迅速扩散到周围环境中,因此它们在植物体内信号的长距离传递中的作用不大。然而,若植物生长在一个密闭的条件下,这些化合物可在植物体内积累并迅速到达作用部位而产生效应。自然条件下发生涝害或淹水时植株体内就经常存在这类信号的传递。 2.化学信号的韧皮部传递 韧皮部是同化物长距离运输的主要途径,也是化学信号长距离传递的主要途径。植物体内许多化学信号物质,如ABA、JA-Me、寡聚半乳糖、水杨酸等都可通过韧皮部途径传递。 一般韧皮部信号传递的速度在0.1~1mm·s-1之间,最高可达4mm·s-1。 3.化学信号的木质部传递 化学信号通过集流的方式在木质部内传递。 近年来这方面研究较多的是植物在受到土壤干旱胁迫时,根系可迅速合成并输出某些信号物质,如ABA。根系合成ABA的量与其受的胁迫程度密切相关。合成的ABA可通过木质部蒸腾流进入叶片,并影响叶片中的ABA浓度,从而抑制叶片的生长和气孔的开放。 4.电信号的传递 植物电波信号的短距离传递需要通过共质体和质外体途径,而长距离传递则是通过维管束。对草本非敏感植物来讲,AP的传播速度在1~20mm·s-1之间;但对敏感植物而言,AP的传播速度高达200mm·s-1。 5.水力学信号的传递 水力学信号是通过植物体内水连续体系中的压力变化来传递的。水连续体系主要是通过木质部系统而贯穿植株的各部分,植物体通过这一连续体系一方面可有效地将水分运往植株的大部分组织,同时也可将水力学信号长距离传递到连续体系中的各部分。 |
| 二、膜上信号的转换 胞间信号从产生位点经长距离传递到达靶细胞,靶细胞首先要能感受信号并将胞外信号转变为胞内信号,然后再启动下游的各种信号转导系统,并对原初信号进行放大以及激活次级信号,最终导致植物的生理生化反应。 (一)受体与信号的感受 受体(receptor)是指在效应器官细胞质膜上能与信号物质特异性结合,并引发产生胞内次级信号的特殊成分。受体可以是蛋白质,也可以是一个酶系。受体和信号物质的结合是细胞感应胞外信号,并将此信号转变为胞内信号的第一步。通常一种类型的受体只能引起一种类型的转导过程,但一种外部信号可同时引起不同类型表面受体的识别反应,从而产生两种或两种以上的信使物质。 一般认为受体存在于质膜上。然而植物细胞具有细胞壁,它可能使某些胞间信号分子不能直达膜外侧,而首先作用于细胞壁。一些外界刺激有可能通过细胞壁—质膜—细胞骨架蛋白变构而引起生理反应。如细胞壁对病原入侵后的防御反应以及对细胞生长发育有重要作用。 (二)G蛋白(G protein) 在受体接受胞间信号分子到产生胞间信号分子之间,往往要进行信号转换,通常认为是通过G蛋白将转换偶联起来,故又称偶联蛋白或信号转换蛋白。G蛋白全称为GTP结合调节蛋白(GTP binding regulatory protein),此类蛋白由于其生理活性有赖于三磷酸鸟苷(GTP)的结合以及具有GTP水解酶的活性而得名。20世纪70年代初在动物细胞中发现了G蛋白的存在,进而证明了G蛋白是细胞膜受体与其所调节的相应生理过程之间的主要信号转导者。G蛋白的信号偶联功能是靠GTP的结合或水解产生的变构作用完成。当G蛋白与受体结合而被激活时,继而触发效应器,把胞间信号转换成胞内信号。而当GTP水解为GDP后,G蛋白就回到原初构象,失去转换信号的功能。 G蛋白的发现是生物学一大成就。吉尔曼(Gilman)与罗德贝尔(Rodbell)因此获得1994年诺贝尔医学生理奖。
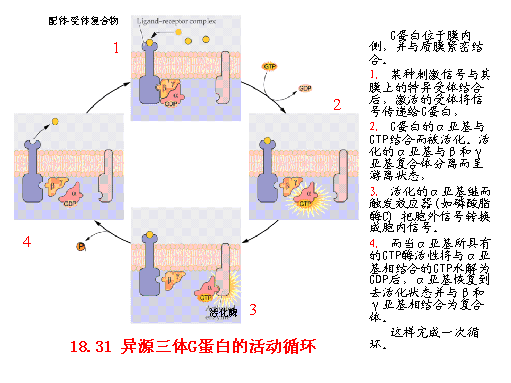
植物G蛋白的研究始于20世纪80年代,并已证明G蛋白在高等植物中的普遍存在,也证明它在光、植物激素对植物的生理效应、在跨膜离子运输、气孔运动、植物形态建成等生理活动的细胞信号转导过程中有重要调节作用。细胞内的G蛋白一般分为两大类:一类是由三种亚基(α、β、γ)构成的异源三体G蛋白,另一类是只含有一个亚基的单体“小G蛋白”。小G蛋白与异源三体G蛋白α亚基有许多相似之处。它们都能结合GTP或GDP,结合了GTP之后都呈活化态,可以启动不同的信号转导。 异源三体G蛋白位于膜内侧,并与质膜紧密结合。当某种刺激信号与其膜上的特异受体结合后,激活的受体将信号传递给G蛋白,G蛋白的α亚基与GTP结合而被活化。活化的α亚基与β和γ亚基复合体分离而呈游离状态,活化的α亚基继而触发效应器,把胞外信号转换成胞内信号。而当α亚基所具有的GTP酶活性将与α亚基相结合的GTP水解为GDP后,α亚基恢复到去活化状态并与β和γ亚基相结合为复合体。如从黄化豌豆幼苗中分离出的G蛋白α亚基的GTP酶可被蓝光激活,说明G蛋白可能参与植物对蓝光的生理过程。利用G蛋白的颉颃剂霍乱毒素(choleratoxin)处理可抑制光敏色素mRNA的积累。说明G蛋白可能参与由光敏色素调节的基因表达及生理过程。赤霉素可引起黄化小麦原生质体的吸胀,而G蛋白抑制剂可消除赤霉素的作用。关于效应器,从已发现的植物第二信使系统来看,主要是cAMP、cGMP、IP3及Ca2 。目前只有PLC(磷酸脂酶C)有充足的证据是G蛋白信号系统中的效应器。近年发现菠菜类囊体蛋白质激酶的活性依赖于由G蛋白激活引起的环腺苷酸(cAMP)浓度升高。随着研究的深入,将发现更多新的植物G蛋白,进一步弄清G蛋白与其受体、效应器之间的作用关系,使对植物体内的信号转导的研究达到新的高度。 |
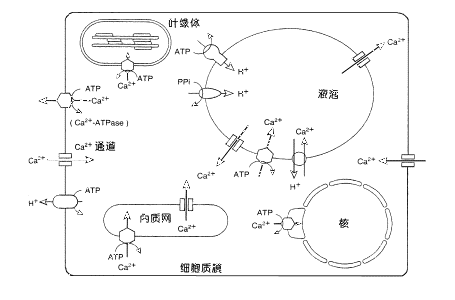
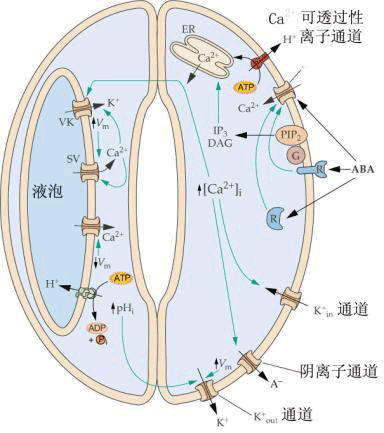
| 三、胞内信号的转导 如果将胞外各种刺激信号作为细胞信号传导过程中的初级信号或第一信使,那么则可以把由胞外刺激信号激活或抑制的、具有生理调节活性的细胞内因子称细胞信号传导过程中的次级信号或第二信使(second messenger)。 1.钙信号系统 植物细胞内的游离钙离子是细胞信号转导过程中重要的第二信使。植物细胞质中Ca2 含量一般在10-7~10-6mol·L-1 ,而胞外Ca2 浓度约为10-4~10-3mol·L-1 ;胞壁是细胞最大的Ca2 库,其浓度可达1~5mol·L-1 ;细胞器的Ca2 浓度也是胞质的几百到上千倍,所以Ca2 在植物细胞中的分布极不平衡。几乎所有的胞外刺激信号(如光照、温度、重力、触摸等物理刺激和各种植物激素、病原菌诱导因子等化学物质)都可能引起胞内游离钙离子浓度的变化,而这种变化的时间、幅度、频率、区域化分布等却不尽相同,所以有可能不同刺激信号的特异性正是靠钙离子浓度变化的不同形式而体现的。胞内游离钙离子浓度的变化可能主要是通过钙离子的跨膜运转或钙的螯合物的调节而实现的,此外,在质膜、液泡膜、内质网膜上都有钙离子泵或钙离子通道的存在(图6-26)。胞外刺激信号可能直接或间接地调节这些钙离子的运输系统,引起胞内游离钙离子浓度变化以至影响细胞的生理生化活动。如保卫细胞质膜上的内向钾离子通道(inward K channel)可被钙离子抑制,而外向钾离子和氯离子通道则可被钙离子激活等。
图 6-26 植物细胞中Ca2 的运输系统 质膜与细胞器膜上的Ca2 泵和Ca2 通道,控制细胞内Ca2 的分布和浓度;胞内外信号可调节这些Ca2 的运输系统,引起Ca2 浓度变化。在液泡内,Ca2 往往和植酸等有机酸结合。 核膜的Ca2 -ATPase在动物中发现,在植物中还不能确认。(前岛正义,1995) 胞内Ca2 信号通过其受体-钙调蛋白转导信号。现在研究得较清楚的植物中的钙调蛋白主要有两种:钙调素与钙依赖型蛋白激酶(后者见本节四)。钙调素(calmodulin,CaM)是最重要的多功能Ca2 信号受体,由148个氨基酸组成的单链的小分子(分子量为17 000~19 000)酸性蛋白。CaM分子有四个Ca2 结合位点。当外界信号刺激引起胞内Ca2 浓度上升到一定阈值后(一般≥10-6mol.L-1), Ca2 与CaM结合,引起CaM构象改变。而活化的CaM又与靶酶结合,使其活化而引起生理反应。目前已知有十多种酶受Ca2 -CaM的调控,如蛋白激酶、NAD激酶、H -ATPase等。在以光敏色素为受体的光信号传导过程中Ca2 -CaM胞内信号起了重要的调节作用。
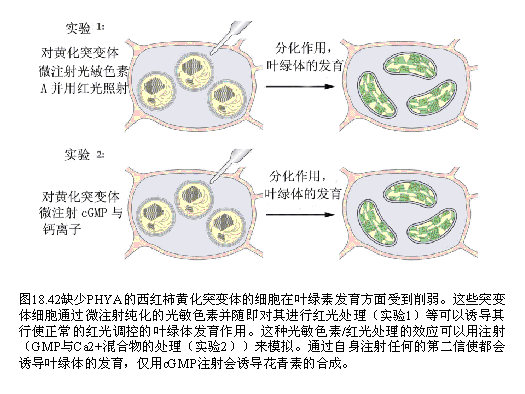
在这个模型中,受体(R)感受ABA的作用导致了Ca2 的输入或Ca2 从内部储存中的释放,从而使得细胞质中的自由Ca2 浓度促进了质膜上阴离子与K Out通道的开放,并抑制了K in通道的开放。当离开细胞的离子比进入细胞的多时,流出细胞水的量就会超过进入的水,如此,细胞就会失去膨胀作用,从而使得气孔关闭。 蔡南海(NamHai Chan)实验室用光敏色素A缺失的番茄单细胞突变系,研究了完整叶绿体中多种光系统色素蛋白质及酶基因表达所必需的光信号传导过程。将信号系统中各组分,如光敏色素A、G蛋白激活剂,Ca2 及CaM、cGMP、cAMP等以及它们的不同组合微注射入单细胞内,随后对叶绿体内多种色素蛋白和酶系在基因活化、转录、翻译三个水平进行观察研究后得出结论:光刺激后,上游的光敏色素与G蛋白激活对完整叶绿体发育及花色素苷合成是共同必须的;Ca2 -CaM系统参与PSⅡ捕光色素、ATP合成酶、Rubisco等光诱导产生,但不参与诱导花色素苷合成,因而只形成不成熟的叶绿体;而在动物视觉系统细胞光感应中起重要作用的cGMP在植物花色素苷诱导中起决定作用,并与Ca2 -CaM一起诱导PSⅠ和Cyt b-6/f的合成。
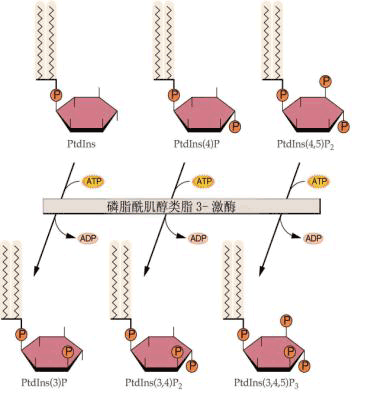
肌醇磷脂(inositol phospholipid)是一类由磷脂酸与几醇结合的脂质化合物,分子中含有甘油、脂酸磷酸和肌醇等基因,其总量约占膜脂总量的1/1O左右。其肌醇分子六碳环上的羟基被不同数目的磷酸酯化 (图6-28),其总量约占膜磷脂总量的1/10左右,其主要以三种形式存在于植物质膜中:即磷脂酰肌醇(phosphatidylinositol,PI)、磷脂酰肌醇-4-磷酸(PIP)和磷脂酰肌醇4,5-二磷酸(PIP2)。
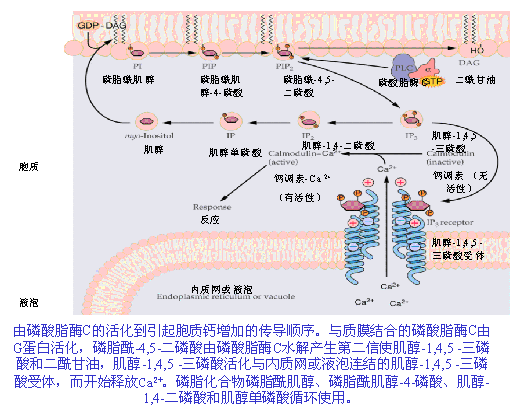
以肌醇磷脂代谢为基础的细胞信号系统,是在胞外信号被膜受体接受后,以G蛋白为中介,由质膜中的磷酸脂酶C(PLC)水解PI2而产生肌醇-1,4,5-三磷酸(inositol 1,4,5-triphosphate IP3)和二酰甘油(diacylglycerol,DG,DAG)两种信号分子。因此,该系统又称双信号系统。在双信号系统中,IP3通过调节Ca2 浓度,而DAG则通过激活蛋白激酶C(PKC)来传递信息(图6-29)。 IP3作为信号分子,在植物中它的主要作用靶为液泡,IP3作用于液泡膜上的受体后,可影响液泡膜形成离子通道,使Ca2 从液泡这一植物细胞中重要的贮钙体中释放出来,引起胞内Ca2 浓度升高,从而启动胞内Ca2 信号系统来调节和控制一系列的生理反应。已有证据证明IP3/ Ca2 系统在干旱和ABA引起的气孔开闭、植物对病原微生物侵染及激发子诱导等环境刺激快速反应中起信号转导作用。 DAG的受体是蛋白激酶C(protein kinase C,PKC),在一般情况下,质膜上不存在自由的DAG。在有DAG、Ca2 时,磷脂与PKC分子相结合,PKC激活,使某些酶类磷酸化,导致细胞反应;当胞外刺激信号消失后,DAG首先从复合物上解离下来,而使酶钝化,与DAG解离后的PKC可以继续存在于膜上或进入细胞质里备用。
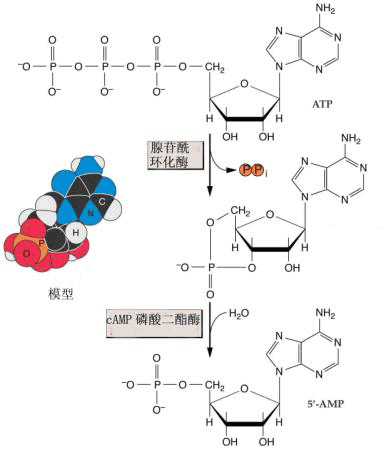
IP3作为信号分子,在植物中它的主要作用靶为液泡,IP3作用于液泡膜上的受体后,可影响液泡膜形成离子通道,使Ca2 从液泡这一植物细胞中重要的贮钙体中释放出来,引起胞内Ca2 浓度升高,从而启动胞内Ca2 信号系统来调节和控制一系列的生理反应。已有证据证明IP3/ Ca2 系统在干旱和ABA引起的气孔开闭、植物对病原微生物侵染及激发子诱导等环境刺激快速反应中起信号转导作用。 DAG的受体是蛋白激酶C(protein kinase C,PKC),在一般情况下,质膜上不存在自由的DAG。在有DAG、Ca2 时,磷脂与PKC分子相结合,PKC激活,使某些酶类磷酸化,导致细胞反应;当胞外刺激信号消失后,DAG首先从复合物上解离下来,而使酶钝化,与DAG解离后的PKC可以继续存在于膜上或进入细胞质里备用。 3.环核苷酸信号系统 受动物细胞信号的启发,人们最先在植物中寻找的胞内信使是环腺苷酸(cyclic AMP,cAMP),但这方面的进展较缓慢,在动物细胞中,cAMP依赖性蛋白激酶(蛋白激酶A,PKA)是cAMP信号系统的作用中心。植物中也可能存在着PKA。蔡南海实验室证实了在叶绿体光诱导花色素苷合成过程中,cAMP参与受体G蛋白之后的下游信号转导过程,环核苷酸信号系统与Ca2 -CaM信号转导系统在合成完整叶绿体过程中协同起作用(图6-27)。
|
| 四、蛋白质的磷酸化和去磷酸化 植物体内许多功能蛋白转录后需经共价修饰才能发挥其生理功能,蛋白质磷酸化就是进行共价修饰的过程,蛋白质磷酸化以及去磷酸化是分别由一组蛋白激酶(protein kinase)和蛋白磷酸酯酶(protein phosphatase)所催化的,它们是上述的几类胞内信使进一步作用的靶酶,也即胞内信号通过调节胞内蛋白质的磷酸化或去磷酸化过程而进一步转导信号。外来信号与相应的受体结合,会导致后者构象发生变化,随后就可通过引起第二信使的释放而作用于蛋白激酶(或磷酸酯酶),或者因有些受体本身就具有蛋白激酶的活性,所以与信号结合后可立即得到激活。蛋白激酶可对其底物蛋白质所特定的氨基酸残基进行磷酸化修饰,从而引起相应的生理反应,以完成信号转导过程。此外,由于蛋白激酶的底物既可以是酶,也可以是转录因子(transcription factors),因而它们既可以直接通过对酶的磷酸化修饰来改变酶的活性,也可以通过修饰转录因子而激活或抑制基因的表达,从而使细胞对外来信号作出相应的反应。 在植物中,目前已知的蛋白激酶至少有30多种,它们的作用也表现在多个方面,包括向光性、抗寒、抗病、根部的向地性、光合作用、自交不亲和性以及细胞分裂等,其中研究得较多的是最初从大豆中得到的依赖于钙离子的蛋白激酶(calciumdependent protein kinase,CDPK),CDPK有一与钙结合位点。现已知的可被CDPK磷酸化的作用靶(或底物分子)有细胞骨架成分、膜运输成分、质膜上的质子ATP酶等。如从燕麦中分离出与质膜成分相结合的CDPK成分可将质膜上的质子ATP酶磷酸化,从而调节跨膜离子的运输。
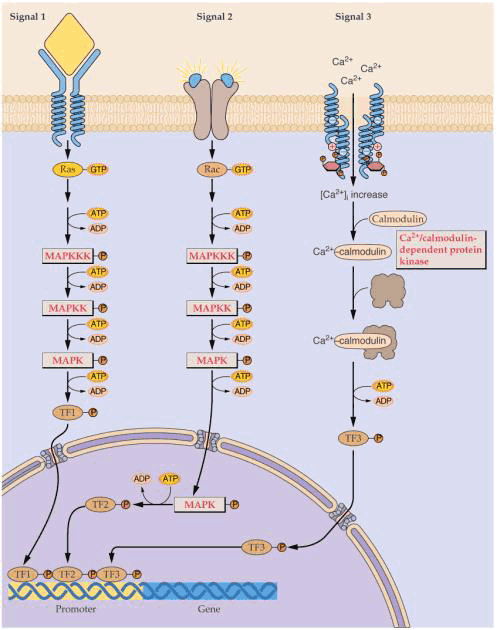
许多由蛋白磷酸化酶传导的信号,级联反应和基因表达。 MAPK-活化的促进细胞分裂的蛋白激酶 TF-转录因子 Ras-一种小的GTP酶 除CDPK类外,还有与光敏色素密切相关的钙依赖型蛋白激酶、同时受钙离子和钙调素调节的蛋白激酶等。近年来在水稻筛管汁液中也测出一个分子量为17 000高度磷酸化的蛋白质以及另一个分子量为65 000,具有蛋白激酶活性的蛋白质,蛋白激酶的活性依赖于Ca2 的存在。筛管汁液内依赖Ca2 的蛋白激酶的发现,说明筛管内存在着信号传导系统。许多植物的SE-CC复合体与周围细胞基本上是共质体隔离的。它一方面在筛管液流中传递着激素等化学信号物质,另一方面又可凭细胞表面质膜上的受体接受外界刺激,进而改变筛管汁液内自由Ca2 的浓度Ca2 作为第二信使激发筛管内的蛋白磷酸化作用,从而调控细胞内一系列生化反应,完成从源到库的信号传导。 蛋白磷酸酯酶逆磷酸化作用,是终止信号或一种逆向调节,与蛋白激酶理论上有同等重要意义,现在植物中鉴定出几种不同的蛋白质磷酸酯酶。如豌豆、胡萝卜中的Type1蛋白磷酸酯酶、豌豆、胡萝卜、小麦中的Type2A和Type2C蛋白磷酸酯酶等。有研究表明,Type1蛋白磷酸酯酶可能与植物细胞的有丝分裂过程的调控有关。在豌豆保卫细胞中存在一种依赖钙离子的蛋白磷酸酯酶,它与K 的转移和气孔的开闭有关。 虽然磷酸化或去磷酸化的过程本身是单一的反应,但多种蛋白质的磷酸化和去磷酸化的结果是不同的,很可能与实现细胞中各种不同刺激信号的转导过程有关。事实上,正是蛋白质磷酸化的可逆性为细胞的信息提供了一种开关作用。在有外来信号刺激的情况下,通过去磷酸化或磷酸化再将之关闭。这就使得细胞能够有效而经济地调控对内外信息的反映。 除上述几类植物细胞信号转导系统以外,还存在其它一些信号转导因子,有人认为,凡是能在胞外刺激信号下发生改变的胞内因子,如果该因子的变化能引起细胞生理活动的变化,则这种因子就是细胞的第二信使。一些化学物质,如H 、H2O2、Mg2 、氧化还原物质以及胞质pH等,是否是植物的胞内信使,尚在探讨之中。此外,以往把信号系统看成是一种因果关系的线性链,然而现在看来,参与传递信号的各因子之间的关系是非常复杂的,信号系统实际是一个信号网络,多种信号分子相互联系,立体交叉,协同作用,实现生物体中的信号传导过程。 |