
第四节 植物亚细胞微结构与功能
植物细胞除原生质膜及外围的细胞壁外,内有众多形状、大小不一的细胞器。它们各自具有特定的生理功能,并协同完成许多复杂的生理过程和代谢反应。此外,在看似无结构的细胞质基质内也进行着多种复杂的反应。
细胞核是细胞遗传与代谢的调控中心,通常将其单独列出介绍,但从某种意义上,细胞核也可看作是最重要的细胞器,故放在此节介绍。
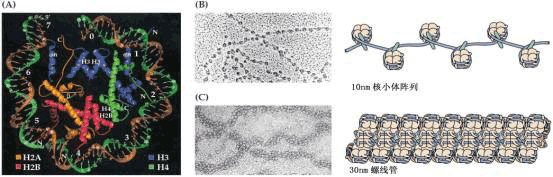
| 一、细胞核 除成熟的筛管细胞外,所有活的植物细胞都有细胞核,其形状与大小因物种和细胞类型而有很大差异。分生组织细胞的核一般呈圆球状,占细胞体积的大部分。在已分化的细胞中,因有中央大液泡,核常呈扁平状,并贴近质膜。 (一)核的化学组成 细胞核主要由核酸和蛋白质组成,并含少量的脂类及无机离子等,其中蛋白质含量最高。在核酸中,DNA含量常高于RNA。核中的蛋白质可分为碱性蛋白和酸性蛋白两类。碱性蛋白质富含精氨酸、赖氨酸,而一般不含色氨酸。它本身带正电,可与带负电的DNA双螺旋结合。染色质的主要组分就是DNA与碱性蛋白质结合形成的核蛋白。与碱性蛋白质结合的DNA不能行使转录功能,即基因被阻遏。酸性蛋白质带负电,富含天门冬氨酸和谷氨酸两种酸性氨基酸,一般还含色氨酸,这可与碱性蛋白质相区别。一般认为酸性蛋白质可解除碱性蛋白质对基因的阻遏作用,而且这种调控具有组织特异性。 DNA在细胞核中的含量是很稳定的,而RNA一般代谢快,其种类和含量有组织差异性。 (二)核的结构和功能 处于分裂间期的细胞核由核膜、染色体、核基质和核仁四部分组成。 1.核膜 核膜(nuclear membrane)由两层单位膜组成。外膜与内质网相连,在朝向胞质的外表面上有核糖体。核膜把核与胞质分隔开,其上有核孔(nuclear pore)。核孔是 由蛋白质构成的复杂结构,叫核孔复合体,它是核质进行物质、信息交换的主要通道。现在认为,核孔复合体是细胞核的独立结构,与核膜仅在结构上有联系。 组蛋白H2A、H2B、H3和H4每种两个结合在一起形成球状结构,盘绕在核粒周围的是DNA。组蛋白H1起连接作用。 2.染色体 染色质(chromatin)是细胞核中能被碱性染料着色的物质,是真核细胞在间期核中的DNA、碱性蛋白、酸性蛋白及少量RNA共同组成的线状复合体。在细胞分裂前,DNA与组蛋白结合,并多次盘绕、超卷曲、折叠形成染色体(chromosome)。染色质和染色体是在细胞周期不同阶段可以互相转变的形态结构。碱性蛋白与DNA形成染色质的基本结构单位——核小体(nucleosome)。每个核小体包括200碱基对(base pair,bp)的DNA片断和8个组蛋白(即碱性蛋白)分子。在核小体的结构中,8分子组蛋白形成紧凑的小圆球,DNA缠绕其上,各个核小体由一段DNA片断(称作连接线)和一个组蛋白分子(H1)相连(图1-9)。整个DNA分子就形成多个核小体相串连的念珠状链。核小体念珠链进一步盘旋、折叠形成染色单体(chromatid)和染色体。从DNA分子到形成染色体的过程中,其长度被压缩了800~1000倍。
A.核小体包括146bp的DNA和由组蛋白构成的八聚体,每个八聚体包含二个H2A、H2B、H3和H4构成的四聚体; B.10nm的核小体阵列; C. 10nm的核小体阵列进一步凝缩成30nm的螺线管结构 3.核基质 核基质(nuclear matrix)是间期细胞核内,除去染色质和核仁之外的非染色或染色很浅的基质。其中除核仁、染色质及核糖体外,含有多种酶。当基质呈凝胶态时称核质(nucleoplasm),呈液态时称核液(karyolymph)。核基质可为核内的代谢提供一个稳定的、良好的环境,为核内物质的运输和可溶性代谢产物提供必要的介质。 4.核仁 细胞核有一到几个核仁(nucleolus),一般呈圆球形,无界膜包围,电镜下可分出颗粒区和纤维区。核仁随细胞分裂周期有消失和重建过程。核仁可合成rRNA和蛋白质。 细胞核是生物遗传物质DNA存在与复制的场所,它控制着基因表达、生物遗传,调节着细胞的代谢、生长与发育。 |
| 二、叶绿体和线粒体 叶绿体(chloroplast)和线粒体(mitochondria)是植物细胞内两类能量转换细胞器,它们的形态特征主要是呈封闭的双层膜结构。尽管它们最初的能量来源不同,但却有着相似的基本结构,且以类似的方式合成ATP。 (一)质体和叶绿体 植物细胞特点之一就是具有双层膜的质体(plastid)。质体是由前质体(proplastid)分化发育而成的。主要有淀粉体、叶绿体和杂色体等。淀粉体(amyloplast)能合成和分解淀粉,内含有一个到几十个淀粉粒,可膨胀得很大。叶绿体含有叶绿素等色素,是光合作用的细胞器,其细微结构、化学组成和生理功能将在“光合作用”一章中作介绍。杂色体可能因含色素的不同而成黄色、橘红色等不同颜色,存在于花瓣、果实、根等各种不同的器官中。 因内外因素的不同,前质体可分化发育成不同的质体。不同的质体之间也可相互转化。如某些根经光照后可以转绿,这就是无色体或杂色体向叶绿体转化的外在表现。当果实成熟时,叶绿体又有可能因叶绿素的退色和类囊体结构的消失而转化为其它有色体。当某种已分化的组织脱分化为分生组织时,某些质体又可回复成前质体。 (二)线粒体 线粒体是进行呼吸作用的细胞器,呈球状、棒状或细丝状等,一般直径为0.5~1.0μm,长2μm左右,不同种类细胞中的线粒体数目相差很大,一般为100~3 000个。通常在代谢旺盛的细胞中线粒体数目较多,反之则较少。如衰老、休眠的细胞或缺氧环境下的细胞,其线粒体数目明显减少。细胞中的线粒体既可随细胞质的运动而运动,也可自主运动移向需要能量的部位。
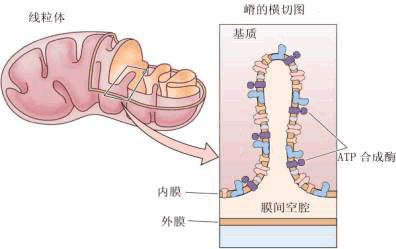
线粒体由内、外两层膜组成。外膜(outer membrane)较光滑,厚度为5~7nm。内膜(inner membrane)厚度也为5~7nm,在许多部位,内膜向中心内陷,形成片状或管状的皱褶(图1-10),这些皱褶被称为嵴(cristae),由于嵴的存在,使内膜的表面积大大增加,有利于呼吸过程中的酶促反应。嵴的数目也可有很大差异,一般需要能量多的细胞,除线粒体数目较多外,嵴的数目也多。在线粒体内膜的内侧表面有许多小而带柄的颗粒,即ATP合成酶复合体,它是合成ATP的场所。线粒体的内膜与外膜在化学成分上不同。外膜磷脂与蛋白质的重量比约为0.82,磷脂多,通透性相对大,有利于线粒体内外物质交流;内膜磷脂与蛋白质重量比约为0.27,为高蛋白质膜,功能较外膜复杂得多,含磷脂少,通透性小,可使酶系统存在于内膜中并保证其代谢正常进行。
线粒体内膜与外膜之间的空隙约为8nm,称为膜间空间(intermembrane space),内含许多可溶性酶底物和辅助因子。内膜的内侧空间充满着透明的胶体状的基质(matrix)。基质的化学成分主要是可溶性蛋白质,还有少量DNA(但和存在于胞核中的DNA不同,它是裸露的,没有结合组蛋白),以及自我繁殖所需的基本组分(包括RNA、DNA聚合酶、RNA聚合酶、核糖体等)。 |

| 三、细胞骨架 细胞骨架(cytoskeleton)是指真核细胞中的蛋白质纤维网架体系,包括微管、微丝和中间纤维等。它们都由蛋白质组成,没有膜的结构,互相联结成立体的网络,也称为细胞内的微梁系统(microtrabecular system)。 细胞骨架不仅在维持细胞形态、保持细胞内部结构的有序性方面起重要作用,而且还与细胞运动、物质运输、能量转换、信息传递、细胞分裂和分化、基因表达等生命活动密切相关。植物的许多生理过程,象极性生长、叶绿体运动、保卫细胞分化、卷须弯曲等也都有细胞骨架的参与。 (一)微管 1.微管的结构 微管(microtubule)是存在于细胞质中的由微管蛋白(tubulin)组装成的中空管状结构。微管粗细均匀,可弯曲,不分支,直径20~27nm,长度变化很大,有的可达数微米。微管的主要结构成分是由α微管蛋白与β微管蛋白构成的异二聚体,这些微管蛋白组成念珠状的原纤丝,由13条原纤丝按行定向平行排列则组成微管(图1-11A)。管壁上生有突起,通过这些突起(或桥)使微管相互联系,或与质膜、核膜、内质网等相连。
A.微管,示13条原纤丝,α、β为微管蛋白 B.微丝。 2.微管的功能 微管的功能是多方面的,主要的功能有: (1)控制细胞分裂和细胞壁的形成 早在进入细胞分裂的前期之前,原来位于周缘胞质中周质的少数微管就被一条环绕整个细胞的,由多个(常多于100个)微管集合成的1~3μm的窄带所取代,由于这条致密的微管带恰好出现在细胞分裂的前期之前,所以被称为早前期带(preprophase band,PPB)。在有丝分裂开始时,此带虽消失,但却在它曾占据的层面留下了某种痕迹,它决定了细胞分裂的部位和分裂面,因为在晚末期形成的细胞板与母细胞壁相结合的位置正是早前期带早先所在的位置。在细胞分裂中,有丝分裂器——纺锤体(spindle)是由微管组成的,它与染色体的着丝点相连,并牵引染色单体移向两极。其后,细胞板的形成与生长也有微管的参与。周质微管决定了纤维素微纤丝在细胞外沉积的走向,在许多不同类型和形状的细胞中,都可见到紧贴质膜之内的微管和紧贴质膜之外的纤维素微纤丝的方向恰好一致,在初生壁、次生壁的沉积过程中,也可见到这一现象。如用秋水仙素(colchicine)等药物处理后,微管解聚,虽不影响新的纤维素微纤丝的产生,但微纤丝排列的模式发生了变化(详见第八章,第二节)。 (2)保持细胞形状 由于微管控制细胞壁的形成,因而它具有保持细胞形态的功能。植物的精细胞常呈纺缍形,这与微管的排列和细胞长轴方向一致有关。当用秋水仙素处理破坏微管,精细胞就变成球形。 (3)参与细胞运动与细胞内物质运输 如纤毛运动、鞭毛运动以及纺锤体和染色体运动都有微管的参与。已经在植物细胞中发现与运动有关的几类微管马达蛋白(microtubule motor protein)。如烟草花粉中的驱动蛋白(kinesin),萱草花粉中的动力蛋白(dynamin),这些微管马达蛋白都与细胞内的物质运输和细胞器的运动直接相关。 (二)微丝 1.微丝的结构 微丝(microfilament)比微管细而长,直径为4~6nm。微丝由收缩蛋白构成(图1-11B),它类似于肌肉中的肌动蛋白,呈丝状,同时还与肌球蛋白、原肌球蛋白等构成复合物质。微丝在植物细胞中有着广泛的分布:通常是成束地存在于细胞的周质中,其走向一般平行于细胞长轴;有的疏散成网状,与微管一起形成一个从核膜到质膜的辐射状网络体系;在早前期微管带、纺锤体及成膜体中也有大量微丝存在。不少实验发现,植物细胞的周质中,微丝与微管平行排列,二者之间还存在相互作用的关系。有实验指出,周质微管的破坏会引起周质微丝的重组,相反,微丝的破坏也会引起微管的重组。 2.微丝的功能 微丝的主要生理功能是为胞质运动提供动力。 (1)参与胞质运动 若从微丝的肌动蛋白(actin)性质及其在胞质中分布的状态来分析,此点不难理解。实验已证明,细胞松驰素B(cytochalasin B)可以使微丝聚集在一起成几十大团,形成网状,如用细胞松驰素B处理轮藻、丽藻等材料,伴随微丝结构的改变,胞质运动也停止。关于胞质运动的机制已基本清楚。大体是微丝中的肌动蛋白与肌球蛋白在胞质内外界面上形成三维的网络体系;肌动蛋白位于外质,肌球蛋白位于内质,肌球蛋白连结着胞质颗粒,在有ATP能量的启动下,肌球蛋白胞质颗粒结合体沿着肌动蛋白微丝束滑动,从而带动整个细胞质的环流。花粉管中原生质流动是肌动蛋白和肌球蛋白相互作用的结果,同时也是花粉管生长的动力。 (2)参与物质运输和细胞感应 已发现微丝可与质膜联结,参与和膜运动有关的一些重要生命活动,如巨噬细胞的吞噬作用,植物生长细胞的胞吐作用。此外,微丝还与胞质物质运输、细胞感应等有关,如中国农业大学阎隆飞等从丝瓜卷须中分离出了肌动蛋白和肌球蛋白,并探明肌动球蛋白是卷须快速弯曲运动的物质基础。 (三)中间纤维 1.中间纤维的结构 20世纪60年代中期,在哺乳动物细胞中发现了10nm粗的纤维,因其直径介于肌粗丝和细丝之间,故被命名为中间纤维,又称中间丝(intermediate filament)。后来在藻类和高等植物中也鉴定出中间纤维。 中间纤维是一类柔韧性很强的蛋白质丝,其成分比微丝和微管复杂,由丝状亚基(fibrous subunits)组成。不同组织中的中间纤维有特异性,其亚基的大小、生化组成变化都很大。 中间纤维蛋白亚基合成后,游离的单体很少,它们首先形成双股超螺旋的二聚体,然后再组装成四聚体,最后组装成为圆柱状的中间纤维(图1-12)。
A两条中间纤维多肽链形成超螺旋二聚体; B两个二聚体反向平行以半交叠方式构成四聚体; C四聚体首尾相连形成原纤维; D 8根原纤维构成圆柱状的10nm纤 维(转引自翟中和,细胞生物学,1995) 2.中间纤维的功能 (1)支架作用 中间纤维可以从核骨架向细胞膜延伸,从而提供了一个起支架作用的细胞质纤维网,可使细胞保持空间上的完整性,并与细胞核定位有关。 (2)参与细胞发育与分化 有人认为中间纤维与细胞发育、分化、mRNA等的运输有关。 |


| 四、细胞内膜系统 内膜是相对质膜而言的,而内膜系统(endomembrane system)通常是指那些处在细胞质中,在结构上连续,功能上相关的,由膜组成的细胞器的总称。主要指核膜、内质网、高尔基体以及高尔基体小泡和液泡等。内膜系统对细胞的生长、分化、成熟具有非常重要的作用。 (一)内质网 1.内质网的结构和类型 内质网(endoplasmic reticulum,ER)是交织分布于细胞质中的膜层系统,通常可占细胞膜系统的一半左右。内质网大部分呈膜片状(图1-13),由两层平行排列的单位膜组成,膜厚约5nm,也有的内质网呈管状,此外,在两层膜空间较宽的地方内质网则呈囊泡状。内质网不是象其它亚细胞组分,只占据细胞中的一定部位,而是能相互连通成网状结构,内与细胞核外被膜相连,外与质膜相连,穿插于整个细胞质中,并且还可通过胞间连丝与邻近细胞的内质网相连。
内质网的形态及数量随细胞类型、代谢活性或发育阶段而异。一般细胞静止期,内质网少,细胞分裂期,内质网增多。内质网本身也在不断更新,是个动态系统。 按内质网膜上有无核糖体的存在把内质网分为两种类型,即粗糙型内质网(rough endoplasmic reticulum,RER)和光滑型内质网(smooth endoplasmic reticulum,SER),前者有核糖体附着,后者没有。粗糙型内质网大多为扁平囊状,靠近细胞核部位。这两种内质网是连续的,并且可以互相转变,如形成层细胞的内质网,冬季是光滑型的,夏季则是粗糙型的。可见,内质网的形态变化是与细胞代谢相适应的。 2.内质网的功能 (1)物质合成 粗糙内质网上的核糖体是蛋白质合成的场所,而光滑内质网参与糖蛋白的寡糖链和脂类的合成。 (2)分隔作用 内质网布满了整个细胞质,将细胞质分隔成许多室,使各种细胞器均处于相对稳定的环境中,有序地进行各自的代谢活动。 (3)运输、贮藏和通讯作用 内质网形成了一个细胞内的运输和贮藏系统。它还可通过胞间连丝,成为细胞之间物质与信息的传递系统。另外,由内质网合成的造壁物质参与了细胞壁的形成。 (二)高尔基体 1.高尔基体的结构 高尔基体(Golgi body)是由膜包围的液囊垛叠而成。液囊呈扁平盘状,囊的两边稍变曲,中央为平板状。通常1个高尔基体由3~12个液囊平叠而成。囊的边缘可分离出许多小泡——高尔基体小泡(图1-14)。
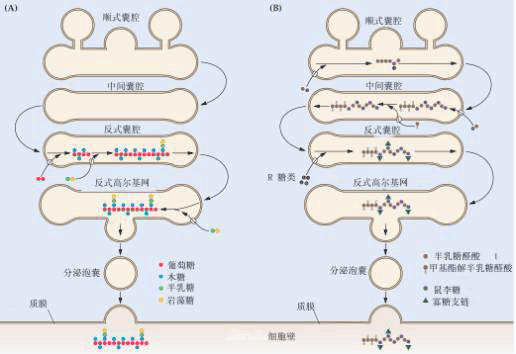
2.高尔基体的功能 (1)物质集运 蛋白质合成后输送到高尔基体暂时贮存、浓缩,然后再送到相关部位。运输的过程可能是:内质网→高尔基体→高尔基体小泡→液泡(分泌液泡)。一些消化酶如α淀粉酶在粗糙型内质网的核糖体上合成后,进入内质网腔,输送至光滑型内质网,然后形成小泡,传送至高尔基体形成面,在高尔基体中蛋白质浓缩成被膜包裹的酶原颗粒(小泡),这些酶原颗粒从高尔基体上脱落下来,最后运至作用部位(参考图1-15)。
A.木糖葡萄糖的集运 B.果胶多糖的集运 (2)生物大分子的装配 高尔基体也参与某些物质的合成或生物大分子的装配。它利用单糖和含硫单糖合成多糖和含硫多糖,是许多多糖生物合成的地点。在合成糖蛋白或糖脂类的碳水化合物侧链时,高尔基体也起一定的作用,很可能糖蛋白中的蛋白质先在核糖体上合成,然后再在高尔基体中把多糖侧链加上去。 (3)参与细胞壁的形成 在植物细胞中,高尔基体的一个重要作用是参与细胞板和细胞壁的形成。例如,组成细胞壁的糖蛋白就是经高尔基体加工,由高尔基体小泡运输到细胞质膜,然后小泡与质膜融合,把内含物释放出来,沉积于细胞壁后形成的(图1-15)。 (4)分泌物质 高尔基体除分泌细胞壁物质外,还分泌多种其它物质。陆生植物根尖最外层的根冠细胞常含许多膨胀的高尔基体。它们分泌多糖粘液,保护并润滑根尖,使之易于穿透土层。食虫植物如茅膏菜和捕虫堇叶腺细胞的高尔基体能分泌破坏寄生组织的酶等。 应提出的是:高尔基体与内质网在功能上具有非常密切的关系,许多生理功能是由二者协同完成的,因此,在细胞中二者常依附在一起。 (三)溶酶体 1.溶酶体的结构 溶酶体(lysosome)是单层膜围绕,内含多种酸性水解酶类的囊泡状细胞器。溶酶体内含有酸性磷酸酶、核糖核酸酶、糖苷酶、蛋白酶和酯酶等几十种酶。 2.溶酶体的功能 (1)消化作用 溶酶体中的水解酶能分解蛋白质、核酸、多糖、脂类以及有机磷酸化合物等,进行细胞内的消化作用。 (2)吞噬作用 溶酶体通过吞噬等方式消化、溶解部分由于损裂而丧失功能的细胞器和其它细胞质颗粒或侵入其体内的细菌、病毒等,所得产物可被再利用。 (3)自溶作用 在细胞分化和衰老过程中,溶酶体可自发破裂,释放出水解酶,把不需要的结构和酶消化掉,这种自溶作用在植物体中是很重要的。例如,许多厚壁组织、导管、管胞成熟时原生质体的分解消化,乳汁管和筛管分子成熟时部分细胞壁的水解以及衰老组织营养物质的再循环等都是细胞的自溶反应。 (四)液泡 1.液泡的结构 液泡是植物细胞特有的,由单层膜包裹的囊泡。它起源于内质网或高尔基体的小泡。在分生组织细胞中液泡较小且分散,随着细胞的生长,这些小液泡融合、增大,最后可形成大的液泡,有的中央液泡(central vacuole)的体积往往占细胞体积的90左右%。细胞质和细胞核则被挤到贴近细胞壁处。 2.液泡的功能 (1)转运物质 液泡借单层的液泡膜(tonoplast)与细胞质相联系。植物细胞利用其液泡转运营养物、代谢物和废物。 (2)吞噬和消化作用 液泡含有多种水解酶,通过吞噬作用,消化分解细胞质中的外来物或衰老的细胞器,起到清洁和再利用作用。 (3)调节细胞水势 大多数植物细胞在生长时主要靠液泡大量地积累水分,并借助膨压导致细胞壁扩张。中央液泡的出现使细胞与外界环境之间构成一个渗透系统,从而可调节细胞的吸水机能,维持细胞的挺度。 (4)吸收和积累物质 液泡可以有选择性地吸收和积累各种溶质,如无机盐、有机酸、氨基酸、糖等。如甜菜根内的蔗糖主要贮存于液泡内;景天科酸代谢植物的叶肉细胞在夜间形成的苹果酸也暂时存于液泡内。液泡膜上存在的质子泵(proton pump,如H -ATPase)可调节细胞内的pH,以维持细胞的正常代谢。液泡内还贮藏一些“代谢废物”或者次生代谢物质,如单宁、色素、生物碱等。 (5)赋予细胞不同颜色 花瓣和果实所呈现的红色或蓝色等,常是花青素所显示的颜色。花青素的颜色随着液泡中细胞液(cell sap)的酸碱性不同而变化,酸性时呈红色,碱性时呈蓝色。在实践中可用花青素的颜色变化作为形态和生理指标。此外,在细胞生长代谢的特定时期,液泡还能形成多种结晶状物。 由上可见,植物液泡不是一个静止的或被动的区域,而是具有多种功能,有明显代谢活性的细胞器。 |


| 五、其他细胞器 (一)微体 1.微体的结构和种类 微体(microbody)外有单层膜包裹,直径为0.2~1.5μm,膜内基质是均一的,或呈颗粒状,无内膜片层结构。根据功能不同,微体可分为过氧化物体和乙醛酸体。通常认为微体起源于内质网。 2.微体的功能 (1)过氧化物体与光呼吸 过氧化物体(peroxisome)含有乙醇酸氧化酶、过氧化氢酶等,所催化的反应参与光呼吸过程。高光呼吸的C3植物叶肉细胞中的过氧化物体较多,而低光呼吸C4植物的过氧化物体大多存在于维管束鞘细胞中。 (2)乙醛酸体与脂类代谢 乙醛酸体(glyoxysome)中含乙醛酸循环酶类、脂肪酰辅酶A合成酶、过氧化氢酶、乙醇酸氧化酶等。其生理功能是糖的异生作用,即从脂肪转变成糖类。 (二)圆球体 1.圆球体的结构 圆球体(spherosome)又叫油体(oil body),是直径约0.4~3μm的球形细胞器,标准的圆球体都含有40%以上的脂类,故也常称其为拟脂体(lipid body)。以前人们认为圆球体是一层膜所构成,但70年代后有人测定了花生子叶的圆球体膜厚度约2.0~3.5nm,只有单位膜厚度的一半,即是由一层磷脂和蛋白质镶嵌而成的半单位膜所组成,磷脂层的疏水基团与内部脂类基质相互作用,而亲水头部基团则面向细胞基质。膜上的蛋白质主要是油体蛋白(oleosins),为圆球体所独有,多为一些低分子量的碱性疏水蛋白。圆球体通常含甘油三酯(triacylglycerols,TAG),存在于谷物的胚和糊粉层、蓖麻种子的胚乳、向日葵与花生的子叶等细胞中。许多油料植物的种子含圆球体特别多。 2.圆球体的功能 种子成熟时圆球体以出芽的方式在粗糙型内质网上形成,其中甘油三酯来自于两层磷脂膜,油体蛋白来自于内质网上的多聚核糖体,形成的圆球体被释放到细胞基质中。当种子萌发时,游离的多聚核糖体合成脂肪酶,经脂解作用的圆球体膜与液泡膜融合在一起,其中的脂肪酸被释放出来参与代谢(图1-16)。圆球体具有溶酶体的某些性质,也含有多种水解酶。
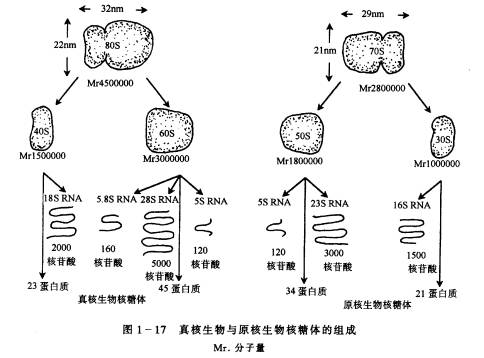
(三)核糖体 核糖体(ribosome)又称核糖核蛋白体,无膜包裹,大致由等量的RNA和蛋白质组成,其多数分布于胞基质中,呈游离状态或附于粗糙型内质网上,少数存在于叶绿体、线粒体及细胞核中。 核糖体由大小两个亚基组成。高等植物细胞质中核糖体的沉降系数为80S,大亚基60S,小亚基40S,大小亚基各由多种蛋白质和相应的rRNA组成(图1-17)。
Mr. 分子量 核糖体是蛋白质生物合成的场所(图1-19)。在一系列复杂的合成过程中,核糖体既要选择所需的各种成分,如对AA-tRNA的选择识别,对多肽链的起始、延长、终止因子的选择识别;又要保持和移动mRNA,这些功能是依靠核糖体中特定部位的蛋白质和rRNA来执行的。 游离于胞基质中的核糖体往往在一条mRNA上成串相连,称为多聚核糖体(polyribosome)。这样一条mRNA链上的信息可以同时用来合成多条同样的多肽链。多种因素(光、生长素、赤霉素、细胞分裂素及细胞发育时期)都可调节多聚核糖体的形成与解聚,从而影响到蛋白质合成的数量。 原核细胞虽然分化简单,但却有核糖体(图1-17),说明了核糖体在细胞生命活动中的重要性。值得注意的是,叶绿体和线粒体内的核糖体大小和分子构成与原核细胞的核糖体更为接近,而与真核细胞基质中的核糖体相差较远。 |

| 六、细胞质基质 在真核细胞的质膜以内,除去可分辨的细胞器以外的胶状物质,称为细胞质基质(cytoplasmic matrix,cytomatrix),也叫细胞浆(cytosol)。细胞质基质是富含蛋白质(酶),具有一定粘度,能流动的透明物质,是细胞重要的结构成分,其体积约占细胞质的一半。很多藻类和高等植物内细胞浆可沿中央液泡环行,被称为胞质环流(cyclosis)。细胞浆还可作穿梭运动(shuttle streaming),其流动方向在一定时间内来回交替。很多代谢反应可在细胞质基质中进行,如糖酵解、磷酸戊糖途径、脂肪酸合成、光合细胞内蔗糖的合成等。细胞质基质还为细胞器的实体完整性提供了所需要的离子环境,供给细胞器行使功能所必需的底物与能量。 综合本节内容,可以看出,植物细胞被内膜系统分隔成多种细胞器,从而使各种生理活动得以按室进行(即代谢、功能的区域化)。然而许多细胞器又因为内膜系统而相互联系,这使得各细胞器之间能进行物质、能量交换与信息传递。应该强调的是,细胞器的分化固然重要,但是,各种细胞器的独立性只是相对的,细胞器离开了完整的细胞虽也能在短时间内进行代谢反应,但不能繁殖和长期生存,而细胞则不然,它可以繁殖并在合适条件下再生出完整的植株。因此,只有细胞才是生物体结构与功能的基本单位。 |
